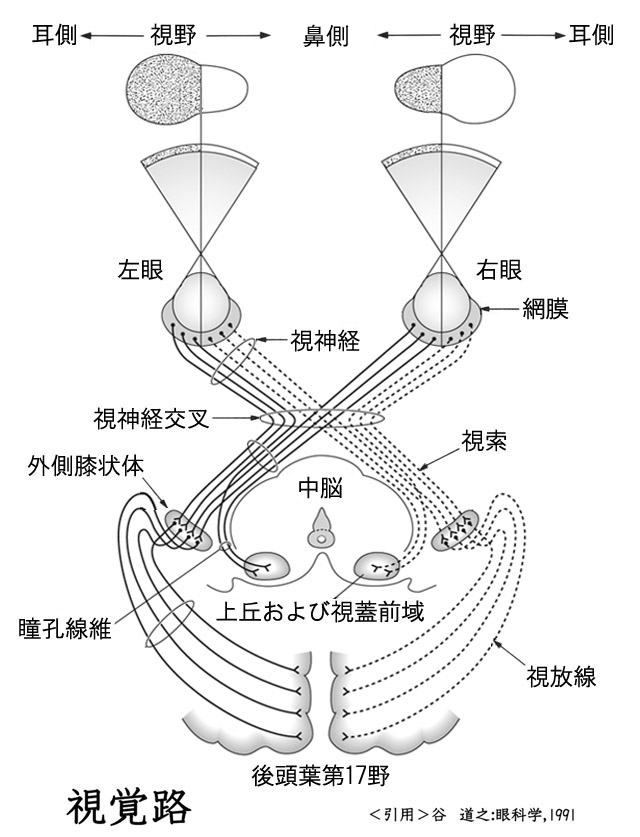
| 視神経・視 路 |
| optic nerve visual pathway |
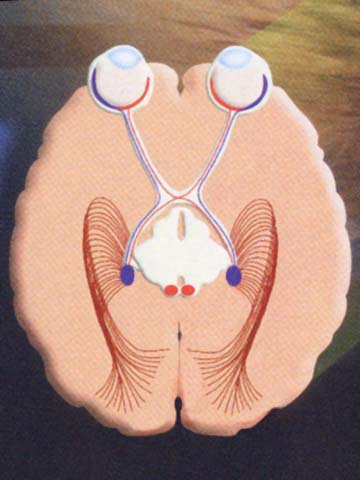
物体からの光は,眼球内で角膜・水晶体の屈折作用を受けた後に眼球の神経層,すなわち網膜に届く.網膜視細胞に与えられた光刺激は符号化(encode)され,視神経を経て大脳後頭葉に達して復号・解読(decode)される.ここで初めて視覚が形成される.視覚とは,光情報による視細胞の活動パターンから生体にとって意味のある情報を取り出す機能である.これらのニューロンの集合である神経経路が視覚路である.
視覚路は神経系の諸経路のなかで最も重要なものに属している.
| 視 路 | |||
| 受容ニューロン | ・・ | 錐体(500万超):明所視・形態覚・色覚 杆体(10,000万超):暗所視・動覚 | |
| ↓ | |||
| 第一ニューロン | ・・ | 双極細胞 | |
| ↓ | |||
| 第二ニューロン | ・・ | 網膜神経節細胞 軸索が視神経線維(100万本超) | |
| ↓ ・・・・・ | ・・ | 視交叉(半交叉) | |
| 第三ニューロン | ・・ | 外側膝状体(視床):中継点 | |
| ↓ ・・・・・ | ・・ | 視放線 | |
| 後頭葉視中枢 → → 高次中枢へ | |||
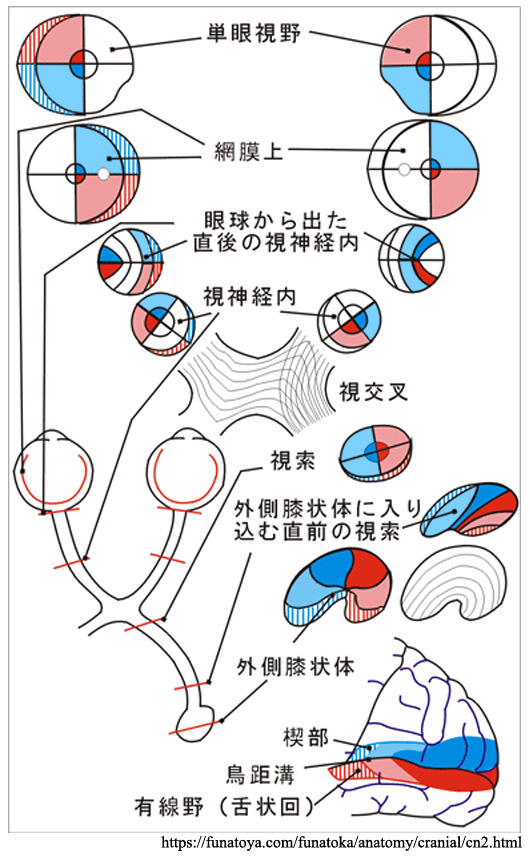
 360°の全視野は,おのおのの眼について耳側視野が網膜の鼻側半分に投影され,鼻側視野が網膜の耳側半分に投影される.
360°の全視野は,おのおのの眼について耳側視野が網膜の鼻側半分に投影され,鼻側視野が網膜の耳側半分に投影される.
これにより,視覚路の障害は視野異常として局在が推定される.
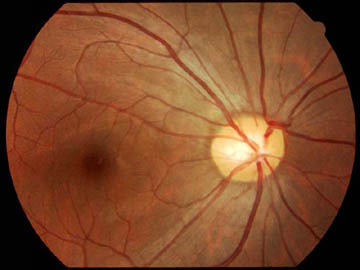
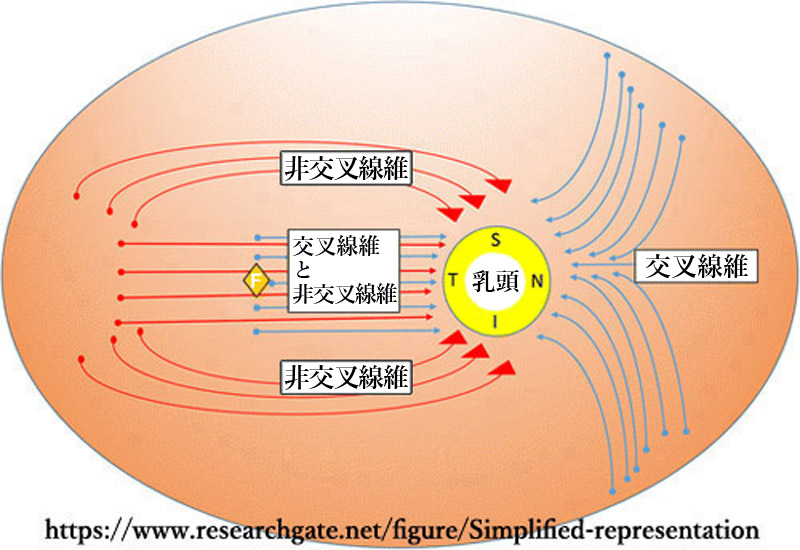
神経線維層 nerve fiber layer
視細胞(受容ニューロン)である錐体および杆体は双極細胞の樹状突起とシナプス結合する.双極細胞(第一ニューロン)の軸索は神経節細胞とシナプス結合する.網膜神経節細胞(第二ニューロン)が出す軸索は網膜の表層で神経線維層となり,後極の一か所;視神経乳頭 optic disk
 |
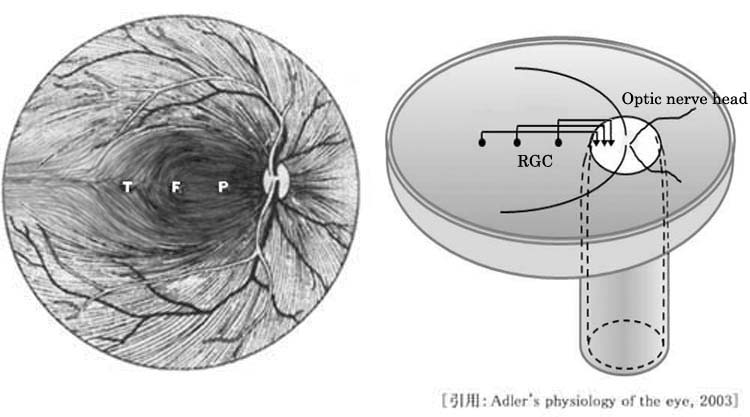 |
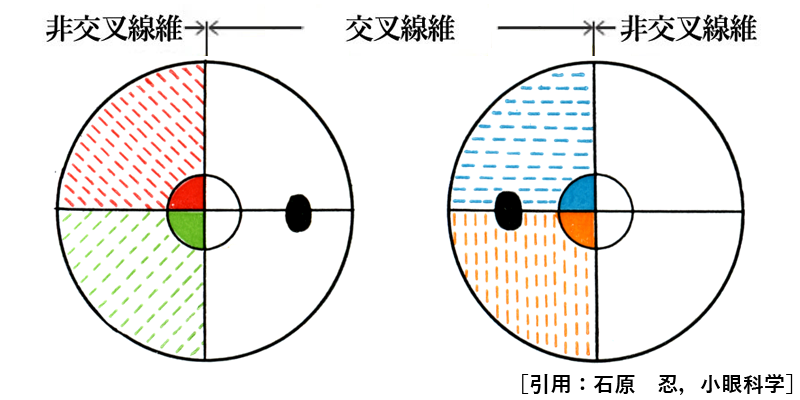
神経線維の走行は特徴的である.模式的には中心窩を通る垂直経線に対し,鼻側網膜の神経線維は交叉線維となり,耳側網膜の神経線維は非交叉線維となる.
➊ 黄斑鼻側線維は,ほぼ直線的に視神経乳頭へ向かい,乳頭黄斑線維 papilloⲻ
❷ 黄斑外側の耳側線維は,中心窩 fovea(上図 F)を通る水平経線で上下に別れ,中心窩周囲を迂回して弓状に走行し,視神経乳頭の上下に集まる.弓状線維 arcuate fibersである.耳側網膜での水平分離線が縫線 temporal raphe(上図 T)である.
❸ 鼻側網膜の神経線維はだいたい直線的に乳頭鼻側へ向かう.放射状線維 radial fibersである.
 |
 |
* 模式的には,これらは交叉線維・非交叉線維に対応する.すなわち,
交叉線維 crossed fibers=➊+❸ ,
非交叉線維 uncrossed fibers=❷ ,と見做すことができる.
【 ☟ 交叉・非交叉 】
 * OCTでの乳頭周囲網膜神経線維層(cpRNFL)厚の計測では,
* OCTでの乳頭周囲網膜神経線維層(cpRNFL)厚の計測では,
視神経乳頭 optic nerve head/optic disk/papilla
視神経乳頭には100〜120万本の神経線維が集合する.乳頭面では,離れた部位からの神経線維は乳頭の外周寄りを,近い部位からの神経線維は乳頭の中心寄りに乳頭内に入る.
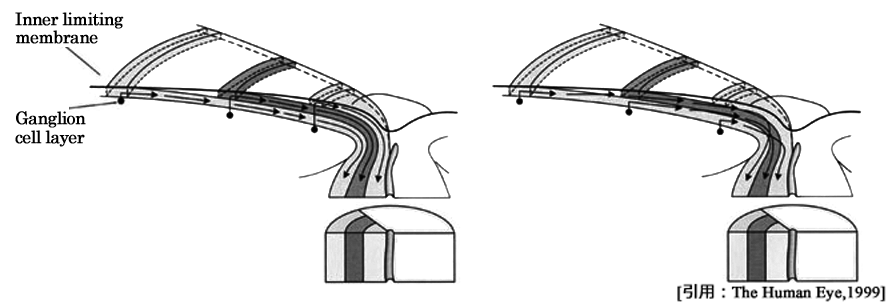
しかし網膜内での神経線維の深浅については,なお議論がある(左図のように考えるのが主流である).
篩状板に入るまでが臨床でいう 乳頭 papillaである.平面的な概念では disk(いわゆる,円板)となる.組織学的には,神経線維が網膜表面から後強膜孔を通過し髄鞘を持つまでの部分が乳頭部となる.
乳頭面に於て網膜内境界膜から連続するのが
*視神経線維の80%は小型神経節細胞からの径1µmの細い線維で,
*眼内の視神経は無髄神経線維である.これにより眼外での伝導速度は高速になるが網膜部では低速である.そんなことで,眼から脳まで信号が伝わるのに0.1秒かかるといわれるほどの遅さである.
*網膜神経節細胞から視神経乳頭までの(伝導)距離は,鼻側網膜すなわち交叉線維のほうが短い.乳頭までの走行が短い鼻側網膜(耳側視野)の情報が,一瞬早く視中枢に届く,とのことである.さらに,中心窩近傍での乳頭黄斑線維束を構成する細い軸索による伝導は最も低速,だそうである.
*視神経は全脳神経の38%という.それにしても 100万本=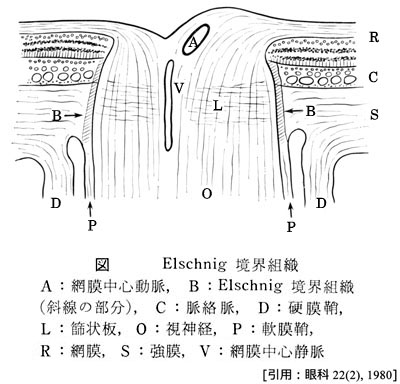
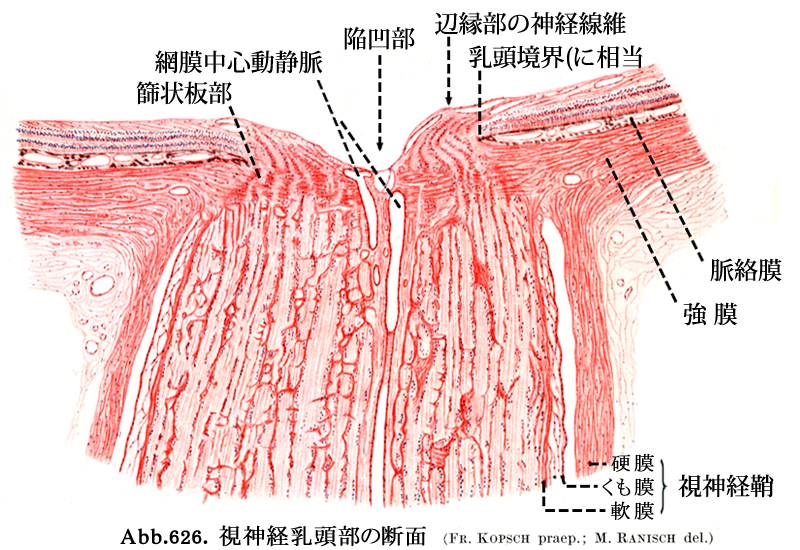
2-1.篩状板部 lamina cribrosa
視神経線維は乳頭内で約1,000本の視束
fascicle
となる.
外周にはグリアを含んだ膠原線維組織が脈絡膜・強膜に対する視神経の境界となっており,
強膜の約半層は篩状板に連続するが,クモ膜下腔の終端になっている.
強膜篩状板を出ると眼外,すなわち有髄の視神経となる.希突起膠細胞
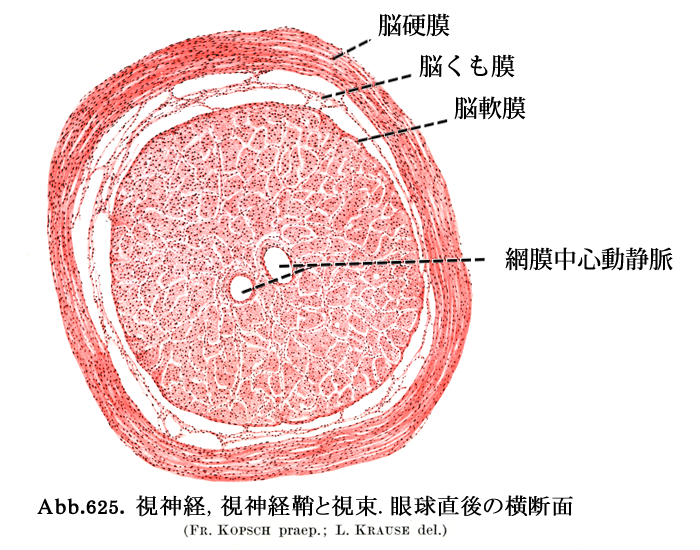
視神経 optic nerve/fasciculus opticus
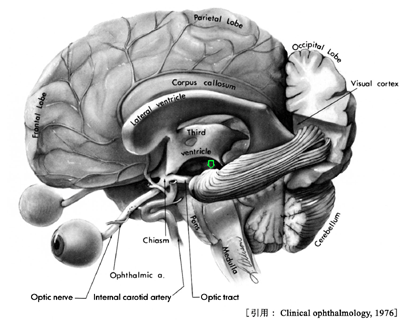
視交叉までが『視神経』である(視路の一部としての名称である).眼外へ出た神経束は強大な神経となり,視神経鞘に包まれて後内側に向かう.神経束そのものの径は約2mm,視神経外径で約3mm.視神経鞘は脳膜の続きであることで,外側から 硬膜・くも膜・軟膜 となる.硬膜鞘を外鞘,くも膜および軟膜を内鞘ともいう.各視束を包むのが軟膜隔壁である.眼球から15〜20mm離れたところで,眼動脈の枝である網膜中心動脈および静脈が外側から入り込む.
*これより中枢の視神経は軟膜の血管網に依存する.中軸近くは末梢に相当し血管支配の条件はよくない,ことになる.乳頭黄斑線維束は中軸部を走行することで不利な位置にある,と説明される.
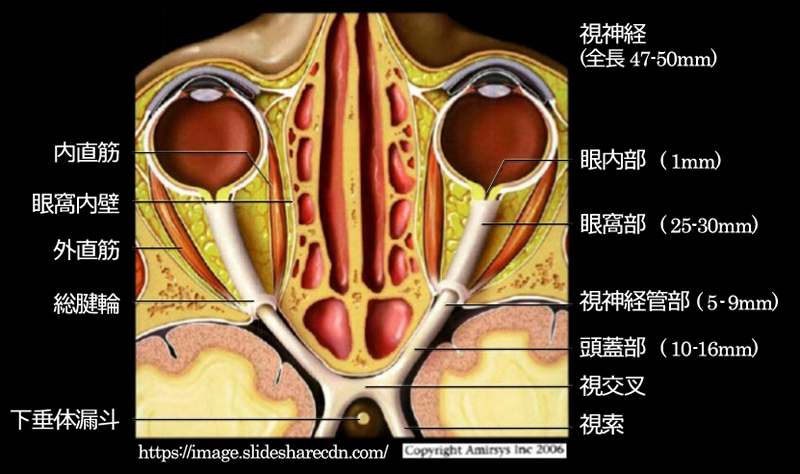 左右両側の視神経は眼窩先端の視神経管を通って頭蓋腔に入り,次第に近づいて脳底部トルコ鞍直上の蝶形骨体・視神経溝で合流して 視交叉 を作る.眼窩内の経路は眼窩先端部まで25〜
左右両側の視神経は眼窩先端の視神経管を通って頭蓋腔に入り,次第に近づいて脳底部トルコ鞍直上の蝶形骨体・視神経溝で合流して 視交叉 を作る.眼窩内の経路は眼窩先端部まで25〜
蝶形骨 トルコ鞍 sella turcica内には下垂体を収納している.
視神経と視交叉との接点を,特に junction(移行部)という.
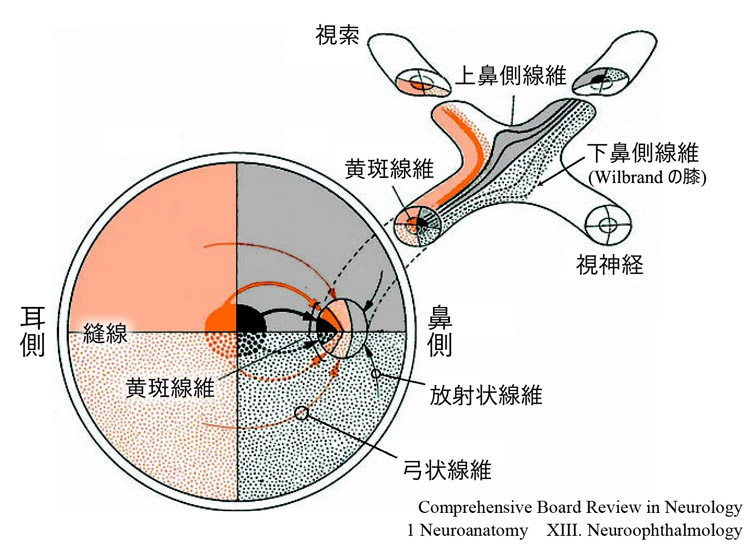
網膜下鼻側の交叉線維は移行部を迂回するような走行で視交叉内に入る.“Wilbrand の膝” である.
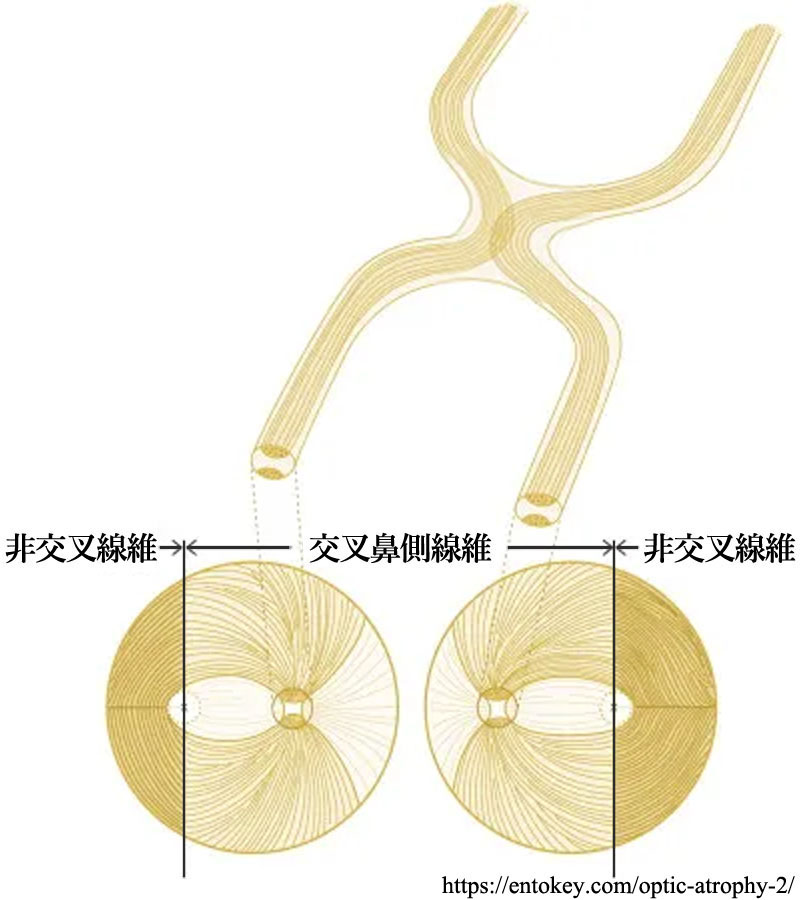
視交叉 optic chiasma と 視索 optic tract
視交叉内では半交叉すなわち,鼻側網膜からの軸索が交叉し対側の耳側網膜からの非交叉性軸索と合流し 視索 を形成する.交叉・非交叉の線維はそれぞれ 約53%・47%,とのことである(Kupfer,1967:要は交叉線維は鼻側網膜由来で面積が耳側より広い).
*交叉線維:中心窩を通る垂直経線の鼻側由来の軸索は,視神経乳頭の耳側に入る乳頭黄斑線維束と乳頭の鼻側に入る放射状線維で構成される.視神経が後方に進むにつれ次第に内側に移動し,対側の視索を経て外側膝状体に至る.交叉線維である.
*非交叉線維:中心窩を通る垂直経線の耳側に位置する軸索は弓状線維として視神経乳頭の上方ないし下方に流入する.視神経の後方では次第に外側に移動し,同側の視索を経て外側膝状体に至る.非交叉線維である.
両眼とも網膜の,例えば右半分は右側の視索にゆく.網膜の右側は視野の左半分に相当するから,右側視索以降では視野の左側半分からの視覚情報が送られている.一方,左側視索は視野の右側半分を受け持っている.すなわち,視索以降の左右の視路では同側眼の耳側網膜由来の視覚情報と対側眼の鼻側網膜由来の視覚情報が伝達される.黄斑を通る垂直子午線部の2°〜数°の範囲は
両側支配(nasoⲻtemporal overlap:鼻側耳側重複)になっていて,左右視野の連続性が保たれるのだとか.
視覚情報は,機能内容によりある程度のまとまりをもって,複数の経路で伝達される.
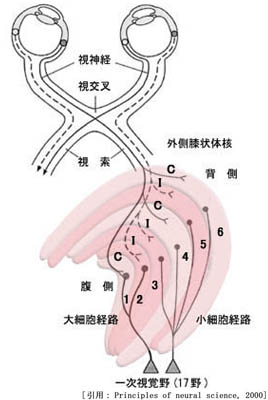
視索部の神経線維は大脳脚の外側を通り,間脳の 外側膝状体 に接続する 膝状体視覚系(retinogeniculate system)と,それ以外の 膝状体外視覚系(extrageniculate system)とに分かれる.前者は皮質経路として物体の形態・色覚・奥行きなどの画像認識に関与し,後者は皮質下経路として瞳孔対光反射のほか視覚性運動反射(追跡)や注視(視覚定位)・空間視・周辺視【視覚チャンネル,明暗チャンネル,定位チャンネル ・・・・・】などに関与する.
神経節細胞以降の信号処理には,それぞれ外側膝状体から視覚野(V1)まで複数の視覚情報の並行する流れがある.
*脊椎動物の進化をたどれば,眼が横を向いていた全交叉から正面を向くようになって非交叉線維が増えてきた,そうである.そういうことで発生学的に,交叉線維は古く,非交叉線維は新しい.
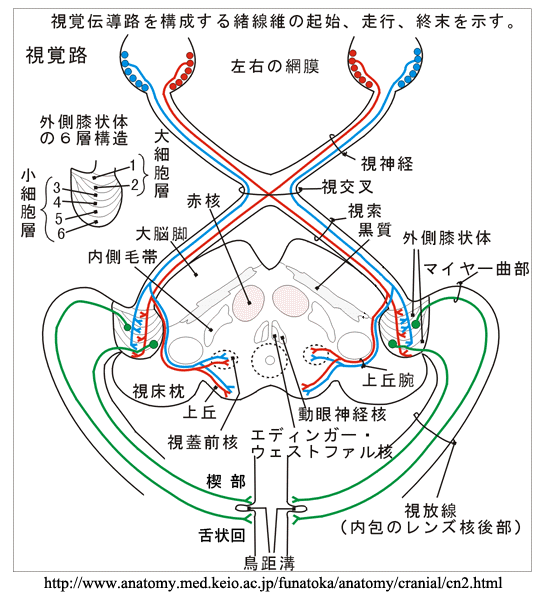
外側膝状体 lateral geniculate body
視神経はここでシナプスを介し,次の第三ニューロンへの信号伝達が行われる.網膜神経節細胞軸索が前部視路となる.
外側膝状体 LGBは,背側部(dorsal,大型)と腹側部(ventral,
階層構造ということで,左右眼の二次元情報から三次元情報を構築している.これにより網膜部位再現地図(retinotopy)が展開されていると考えられている.視床の神経核の一.
網膜上半部(1∕4)由来の線維(下部視野)は外側膝状体の内側に届く.
網膜下半部(1∕4)由来の線維(上部視野)は外側膝状体の外側に届く.
網膜外の入力源:下記
C;contra-lateral:contra = "against","opposite"
I;ipsi-lateral:ipse = "self"
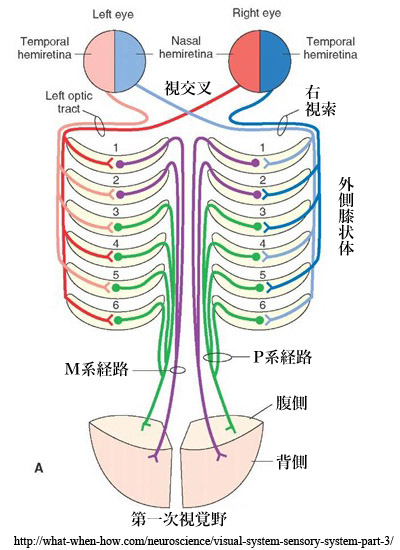
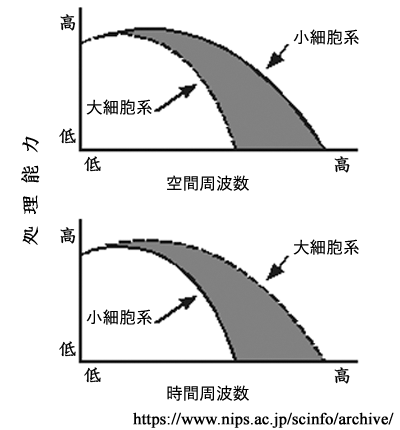
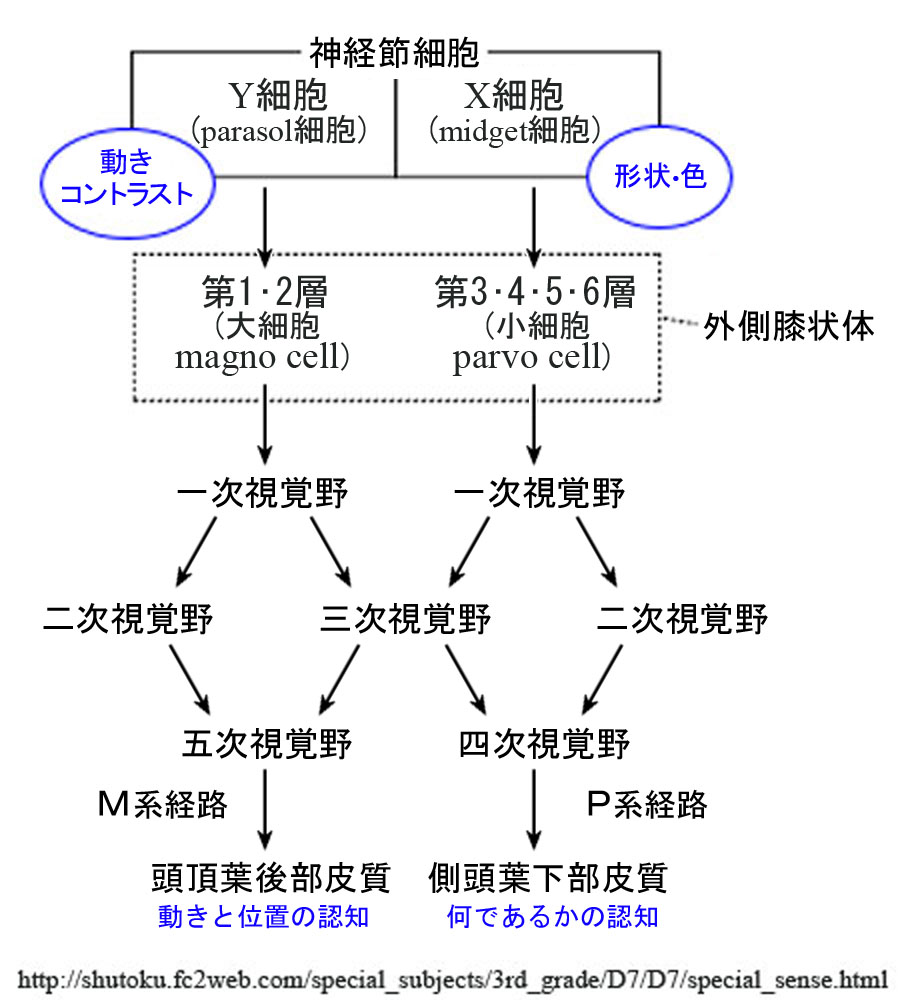
●M系( magno cell system:大細胞系) と P系( parvo cell system:小細胞系)
外側膝状体の中継細胞には,おおきくM系とP系の二種類の細胞群がある.
これにより,網膜から外側膝状体を経て一次視覚野に至る経路は,
大細胞系経路(magno 系)と小細胞系経路(parvo 系)とに分けて説明される.
①M系(magnocellular system:大細胞系):
錐体・杆体(網膜全体)からの入力を受けたY型(parasol cell,α細胞,大型)神経節細胞の信号は大細胞層(1,2 層)で中継される.受容野は広く低分解能(低い空間周波数)であるが一過性反応・高感度(高い時間周波数)で,動きの検知(運動視,追視)・立体視(空間視)などに関わる.全軸索の10~15%ほどで,主に運動処理として背側経路の情報になる.色覚の情報はない.
②P系(parvocellular system:小細胞系):
錐体(主に中心部)からの入力を受けたX型(midget cell,β細胞,小型)神経節細胞の信号は小細胞層(3,4,5,6 層)で中継される.受容野が小さく持続性反応・低感度(低い時間周波数)であるが,高分解能(高い空間周波数)で形態覚や色覚(red−
両系は網膜内では混在しているが,外側膝状体では明確に分離され層状に配列・分布する.
高次中枢皮質の上で大細胞系の情報と小細胞層の情報の収束が起きると考えられている.
| 【大細胞系経路】 | 【小細胞系経路】 | |
| 神経節細胞 | 大型 | 小型 |
| 空間分解能 | 低い | 高い |
| 時間分解能 | 高い | 低い |
| コントラスト感度 | 低周波域 | 高周波域 |
| 波長選択性 | ない | ある |
| V1野 終端層 | 4Cα | 4Cβ |
【 ☞ 時間周波数・空間周波数は こちら 】
③顆粒細胞層(koniocellular layer):
W型(γ細胞,bistratified)神経節細胞からの神経線維は層外の顆粒細胞(K細胞)に接続する.
P系からの入力もあり,色覚(blue᠆yellowチャンネル)を担うらしい.K層はM1腹側のK1からP3-4間のK6まで存在する.
➊ Mは,神経節細胞では『midget』の,外側膝状体では『magno』の略として,Pは,神経節細胞では
これにより混乱するが,
大細胞M系 ← Y型神経節細胞(parasolまたはPα),
小細胞P系 ← X型神経節細胞(midgetまたはPβ),となる.(ここの第3の P は primate,神経節細胞は頭文字には略さない).
なおX型・Y型ともサルのM系に対応し,P系は高空間解像度と色覚のためにある,と解釈されている.また,色覚のメインはK系ともいわれている.
➋ 網膜神経節細胞の復習は 第四章 で.
➌ 高次視覚野への経路は 第十章 で.
 |
 |
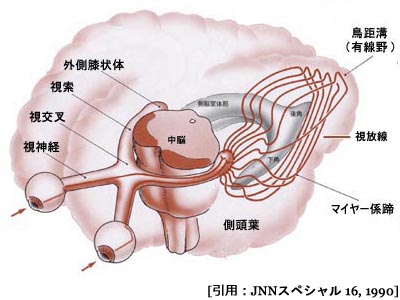
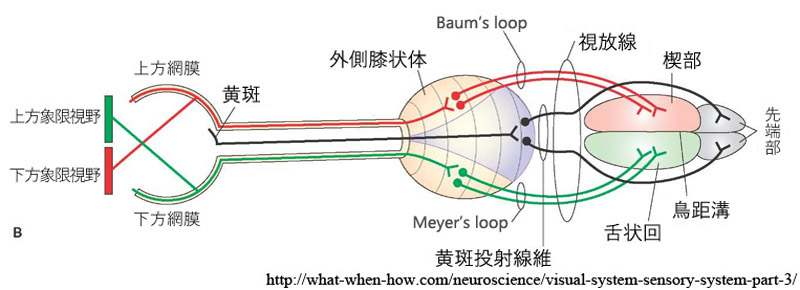
視放線 optic radiations
外側膝状体(上右図,緑矢印部が外側膝状体)から始まる第三ニューロンの軸索線維は側頭葉の内側面,内包といわれる線維群の後外側面から発し,やや前方に進んだ後に後頭葉へと向かう.膝状体鳥距路,すなわち 視放線 optic radiations
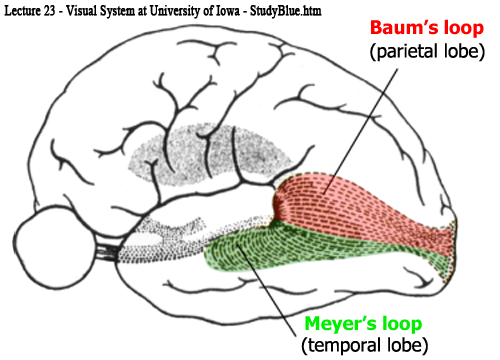
網膜上半部由来の線維(下半視野)は(背側部から)上方へ広がり頭頂視放線となり,頭頂葉 parietal lobeを通過して後方の
網膜下半部由来の線維(上半視野)は(腹側部から)前外方へ広がり側頭視放線となり,側頭葉 temporal lobe・側脳室下角を包むようにして周り下角及び後角の外表面から下鳥距皮質・舌状回に達する(Meyer係蹄 またはArchambault).
LGB の中心部からおこる黄斑線維は視放線の広い中心部を占めるほか,視放線内の線維の配列も多少の混合があるが局在の原則は保たれている.
 |
 |
第三(外側膝状体)ニューロンの軸索は,視放線 optic radiationsあるいは外側膝状体鳥距溝路線維(膝状体有線領経路)
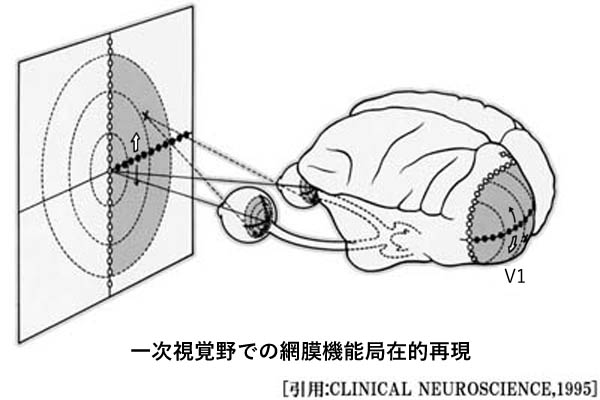
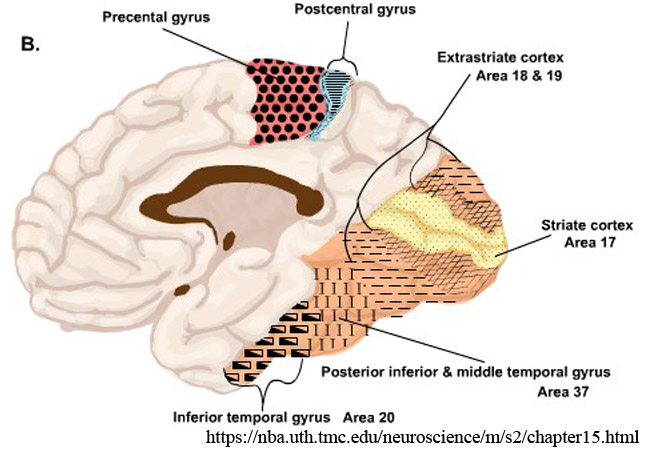
一次視覚野 primary visual areaは,V1野,Brodmann area 17,17野,有線領,線条野,などと呼ばれる.
 |
 |
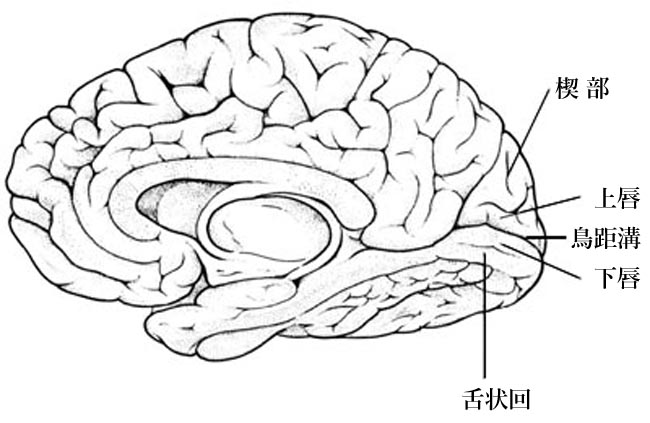
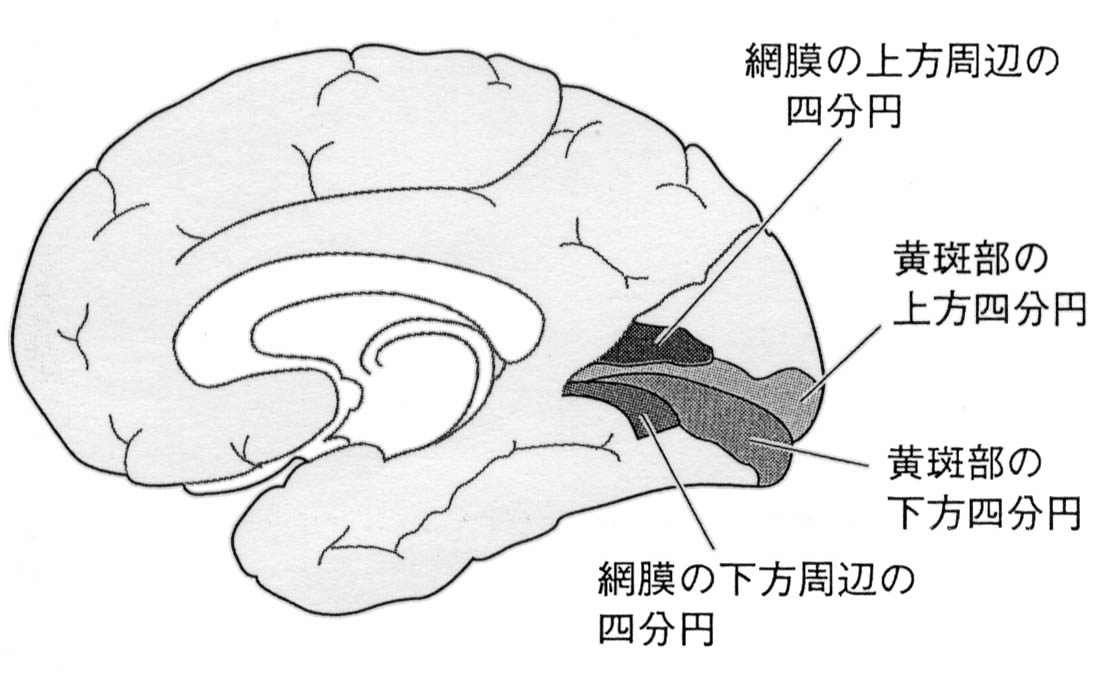
視覚野皮質は後頭極より始まり,楔部と舌状回にはさまれた範囲を示す.ここに対応する網膜部位の視覚印象が投射される(Henschen の『皮質網膜retinotopy』).上下視野の区別は視覚伝導路の全体で維持されており,網膜の上部(下方視野)は鳥距溝の上方(上壁あるいは上唇)に,下部(上方視野)は下方(下壁・下唇)に再現される.
 |
 |
中心窩の部分(再現域)は特に皮質の大きな部位を占め,かつ両側支配と考えられている 網膜での乳頭黄斑線維束は明らかに耳側視野すなわち交叉線維であるが,中心窩の上下線維は両側に振り分けられるという.また,中心窩線維は視神経内で再配列される.
黄斑領域の線条体全域に対する割合は,網膜の中での黄斑部の割合よりも相対的にはるかに大であることは,網膜の中で黄斑は感覚細胞が最も密な部分であること,視力 visual acuityが最も高度であるという事実に一致する.
【 ☟ 皮質拡大機能 】
| 後頭葉視覚中枢(鳥距溝)と視野との関係(Holmes) | 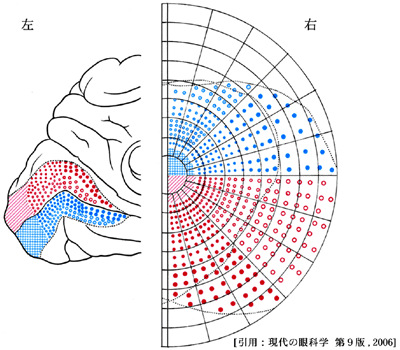 |
| 鳥距溝において網膜の黄斑領域は比較的広い範囲に, 網膜の周辺領域は狭い範囲に投射されている. 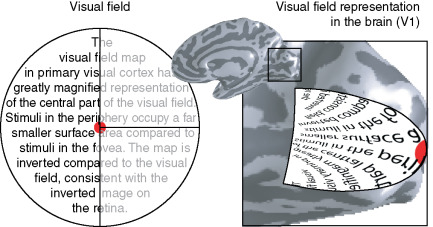
|

線条野の最前域は網膜の鼻側(内側)に対応し,周辺視野すなわち単眼視視野である最も外側(耳側)の半月部 crescentに相当している.
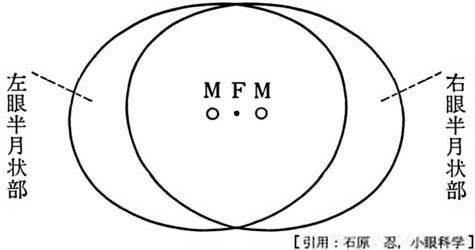
最前域とは深部・内側の第三脳室寄りのこと.右図で 1-2 と 8-9 がそれぞれ左眼・右眼で単眼視している部分を示している.
 大脳で視覚情報を処理しているのが視覚領で,30何か所の視覚野がある.
大脳で視覚情報を処理しているのが視覚領で,30何か所の視覚野がある.
網膜は視覚情報を点として認識し視覚野に伝達する.脳では,V1野 において点を線に構成する.すなわち,視覚情報の輪郭・奥行き・動き等の特徴抽出が始まる.V2野 は視覚連合野としての初めの領域で,線を形に構成する.外側膝状体すべてのサイズの細胞の信号は V1野 へ行く.大細胞は V2野 へも投影される.
機能的に V2野 は V1野 と多くの共通な特性を持ち,V1 と V2 には,網膜上の位置・奥行き・動き・形・色などの視覚情報のすべてがほぼ平等に伝達される,とのことである.これにより一次二次視覚野は “低次視覚野” とも云われる.
これ以降の視覚情報処理は形態情報と空間情報の大きく二つのルートで行われる.
形,色などは VP野(ventral posterior area,腹側後部領域,腹側三次視覚野),四次視覚野を経て側頭連合野へ送られる(なに情報).
位置,奥行き,動きなどは 三次視覚野,MT野(middle temporal area,五次視覚野)を経て頭頂連合野へ送られる(どこ情報).
高次視覚野
に情報が送られると,物体の形の認識,奥行きの認識,運動の認識,色の認識(色覚関連中枢は後頭葉底部の舌状回と紡錘状回に存在する)などが可能となる.
● 以上のように,視覚とは『見ているものはなにか』『どこにあるか』と『視線の移動』によって成り立つといえる.
● 有線領,線条野 striate cortex:Gennari/Vicq d'Azyr
よく発達した顆粒層 granular cortex と線維のため肉眼でも白く線状(層構造)に見える.第4層に顕著.
後頭葉の内側面後部にある鳥距溝をはさむ皮質のこと.
● calcarine fissure の日本語は ?
いくつかの教科書を見渡すと,誤植?と思われるような表記に出会う.うかつであった.何と,本邦近代眼科の始祖「小眼科學(石原 忍〜大正14年1925年創著)」のとある版では“烏距溝”(烏〜カラスです)とあるではないか,webでも,検索してみると混乱しているようである.なんたることか.ウキョコウ ? フォントの所為 ? ウン十年付き合っている私の版では「トリ」になっている.やれやれ.
calcarine;calcar avis=けづめ;蹴爪;距,ということで,カラスの足なのでしょうか,単なるトリの足なのでしょうか.
● 外側膝状体・上丘を第一次視覚中枢として,後頭葉を第二次視覚中枢という分けかたもある.
そんな訳で視覚求心路とは,視神経線維の終了する外側膝状体・上丘・視蓋前域までを指す.
●
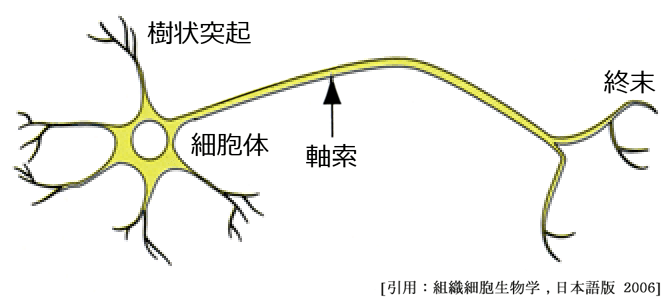
神経細胞(neuron):神経系を構成する基本単位.
【 ☞☞ こちら で 】
●
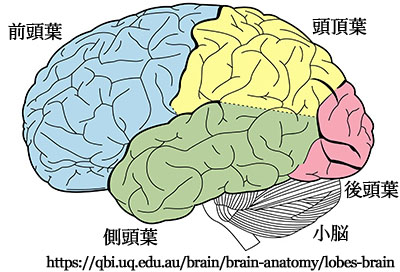 大脳皮質の区分:
【 ☞☞ こちら で 】
大脳皮質の区分:
【 ☞☞ こちら で 】
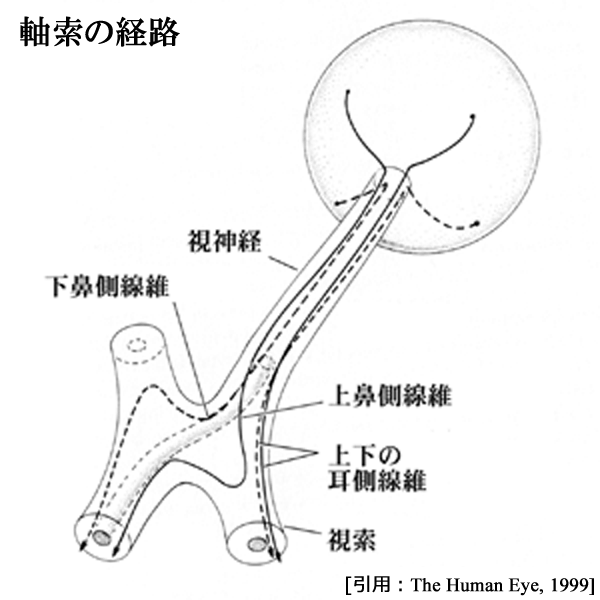
● Wilbrand's knee と junction scotoma
視交叉では,両眼の鼻側から来る線維は交叉し,それぞれ反対側の視索へ向かう.このとき,網膜下鼻側象限からの線維(右図破線)は,視交叉前角をいったん反対側の視神経視交叉接合部を前進し大回り(前脚╱Wilbrandの膝)してから交叉する.ただし人工産物であるとして 否定的な研究者 もいる.
交叉する時上下線維はバラける,のは確からしい.網膜上鼻側象限からの線維(右図実線)は視交叉後角を廻って交叉する.後脚である.
Wilbrand線維と黄斑線維は接近しており,視交叉前部の障害では,同側の(半盲性)中心暗点と他眼の耳側上方の周辺視野欠損を特徴とする.junction scotoma
☑ Prof. Hermann Wilbrand (1851᠆1935) Hamburg, Deutschland:ophthalmologist.神経眼科の先生.
間違え易いのは
☑ Prof. Erik Adolf von Willebrand (1870᠆1949) Helsinki, Finland:internist.血液学の先生.
英語読みでは「ヴァン・ウィルブランド」,ドイツ語読みでは「フォン・ヴィレブラント」,フィンランド語読みでは「ヴォン・ヴィレブランド」,だそうです.
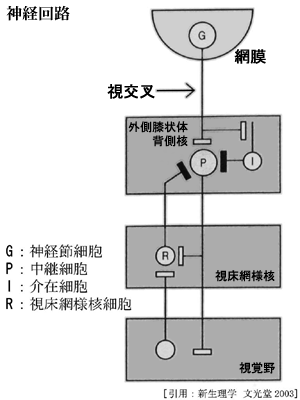
●外側膝状体:LGB 【 return 】
・LGB d(背側核).大脳皮質へ視覚情報を伝える.
・LGB v(腹側核,あるいは膝状体前核)は,視神経線維,視覚野および上丘から視覚入力を受け,共同眼球運動の制御をしている.上丘 superior colliculus,視蓋前域 pretectum,視交叉上核 suprachiasmatic nucleus,不確帯 zona incerta,副視索の外側終止核 lateral terminal nucleus of accessory optic tract,などの皮質下核に線維を送る.さらに小脳への連絡もある.
・大脳皮質の視覚領野・視床網様核・脳幹網様体からの入力 ・・・・・
LGB の活動は後頭葉皮質により影響されうる(皮質膝状体線維).視覚インパルスの中枢性コントロールは介在性ニューロンによる抑制をうけ,選択的注意 selective attentionにとって重要とされる.
1/4は抑制性介在ニューロンでコントラストの増強しているという.
網膜からの興奮性シナプス入力は 1/5程とのことである.
これらにより,左右眼の入力に対しバランスをとるようなある種の関門作用の部位とも説明される.
●膝状体外系 extrageniculate system 非膝状体系 【 return 】
視神経の一部(基本的にY細胞系の約1% (?)と,W細胞系)は外側膝状体背側核に入らず,上丘・視蓋前域核・視交叉上核・視床枕核などへ連絡する.これら膝状体を経由しない入力が外側膝状体外系である.
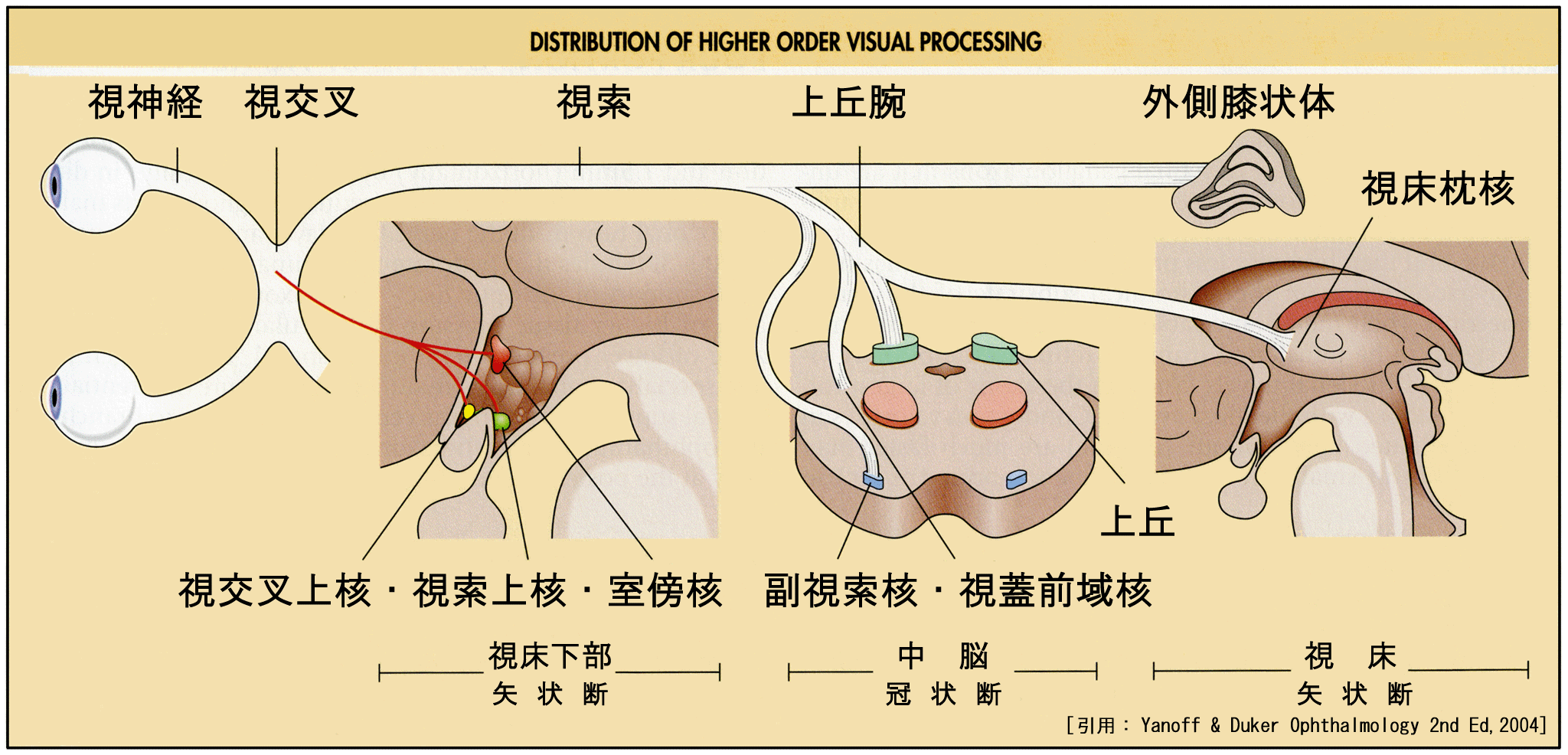
例えば非膝状体系の代表は,上丘(retinocollicular投射)・視蓋前域(retinopretectal投射)へ向かう
視覚反射
の経路である.上丘は進化の中で原始的な視覚入力を受ける部位で,視蓋tectumという.これより前方部が視蓋前域となる.
哺乳類以外の脊椎動物において,中脳の視蓋optic tectumは主要な投射部位で,副視索核を含む.
tectum vision は
盲視
に関係する.また,膝状体系の未成熟な新生児期においては,重要な視覚経路とされる.
●上丘 superior colliculus :
中脳 midbrain ∕ mesencephalon にあり,空間位置情報を処理し眼球運動や頭部運動の制御をしている.
視覚や音の空間的地図や体表面の地図がある.これにより,視環境の中で対象への視線を定位する(見たい対象に顔を向ける)こと(注視 foveation)や,saccade運動に関連する視運動反射(眼-頭位反射,追跡・追従運動)の中枢である.
| 表層3層 | … | Y細胞・W細胞と大脳皮質視覚野からの入力を受け視覚処理を行う.網膜再現地図がある. |
| 中間層 | … | 眼球運動地図.視覚対象の位置を測っていると考えられ,saccade運動に関連する. |
| 深層3層 | … | 視覚系,体性感覚,聴覚,前庭感覚系からの入力を受け,脊髄や延髄に出力する.ということで, 頭部の位置補正など視覚情報に対する反射的姿勢運動に関連し,空間定位機能を統合していると考えられる. |
★入力線維:入力信号の最大の発信源は前頭眼野とのことである
① 網膜視蓋(上丘)路 retinotectal tract:網膜より視神経を通って上丘に至る線維.Y型(α細胞,
視神経は視交叉にて半交叉するので一側の視神経は両側の上丘あるいは視蓋前域に投射する.
② 上丘へは外側膝状体を経由して入力 ?
③ 皮質視蓋路 corticotectal tract:前頭眼野や大脳皮質視覚野より起こり同側の上丘あるいは視蓋前域に至る.
★出力線維:視床枕を経由して高次視覚野に投射する
情報の解析には,第2次視覚野,頭頂葉などへ行く.
眼球運動信号は,脳幹網様体へ行く.ここから運動神経核が制御される.
●視蓋前域核 pretectal region(視蓋前野:pretectum) :
上丘のすぐ前方の位置で脳幹と間脳の境界にあり,瞳孔調節や視運動性眼球運動などの反射中枢.
視索核・視索前域オリーブ核・視蓋前域前核・視蓋前域後核などの神経核の複合体で 視蓋前域のほぼ全域に網膜からの線維がシナプスを作り明るさの変化を伝えている(明るさの変化,すなわち瞳孔反応と関連).
W型(γ細胞,小型)・X型(β細胞,midget cell,中型)神経節細胞からの軸索のほか,外側膝状体腹側核や上丘からの線維が終止する(すなわち調節反応と関連).
遠心性線維はE-W核のほか,外側膝状体腹側核・外側膝状体背側核・視床の後外側核などと連絡する.
●視交叉上核 suprachiasmatic nuclei :
メラノプシン神経節細胞からの投射を受け 概日周期 の調節
(24時間周期に同調)
をする.
視交叉付近の表面線維が連絡しているらしい.
下垂体ホルモン・松果体ホルモンの調節に関与する(神経内分泌調節).
●視床枕核 pulvinar :
眼球運動に関連する制御らしい.
上丘や視蓋前域から入力を受け,高次視覚野(特に頭頂連合野)に投射する.
●副視索核 nucleus of the accessory:
副視索路 accessory optic tract は,反射性の眼球運動や瞳孔反射など画像認知に直接関係しない視覚信号を送る経路.W細胞からの投射を受ける.小脳と連絡し,自身の位置情報を基に網膜像の動きを検出し,網膜像を安定させている(ブレ抑止)らしい.
●終核(終止核) terminal nucleus :
前庭核と小脳へ連絡し 眼位と頭位を統合する副視索路
内側終止核・背側終止核・外側終止核
●視覚反射
視覚反射には,瞳孔反射・調節反射のほか眼球運動に関する追従・固視が含まれる.
大脳皮質の関与しない皮質下反射は,おもに上丘が関わる注視反射と,視蓋前域が関わる対光反射がある.注視反射は眼球の方向・位置を調整し,対光反射は光量を調整する.
大脳皮質が関与する皮質性反射は,おもに後頭葉が関わる注視反射と,視蓋前域が関わる調節反射がある.注視反射は眼球の方向・位置を固定し,調節反射は焦点を対象に合わせる.
●視床
thalamus :
すべての感覚の伝導路は視床を通る.視床網様核は視床の全般的な働きを抑制する.
松果体がある.その昔,生物の進化の初期の段階で原始的な視覚を担っていたとのことで,松果体は第1の眼と云われる.その後に脳が左右ペアとして発達することで二つの眼が別の位置に進化することになり,松果体は違う役に代わったのだとか.
●サーカディアンリズム circadian rhythm:
人間の体は,25時間の周期を持って活動や睡眠をとったり,ホルモンの分泌等を調節している.この周期(サーカディアンリズム・概日リズム・生物時計)は視床下部の視交叉上核(SCN)によってコントロールされている.神経節細胞にあるメラノプシン(melanopsin)という光受容タンパク質が網膜に入る外界の光を感じて SCN にシグナルを送りサーカディアンリズムと昼夜のリズムを同期化しているといわれる.この調節には杆体や錐体からも信号が届いている.情報の一部は
松果体
に伝わり,明るいときには
セロトニン(延髄の縫線核が分泌)
暗いときには メラトニン(松果体が分泌)により,生体にリズムを与え実時間に合う活動ができている.
メラトニンを投与すると体内時計が進む.ある種の睡眠障害や海外旅行での時差ボケは,昼夜のリズムが実時間に同期しないために起こる.従って,光暴露によってメラトニン量を減らし時計をリセットすることが必要となっている.視力喪失者も同様に体内時計を合わせる必要がある,とのことである.光暴露〜光感受性細胞は体中に分布しているそうである.
🗸ブルーカット : メラノプシンは青色光(400〜500nm)感受性があり,体内時計には青色光が必須となっている.昼の活動に必要な青色光でも,夜間では睡眠の質を悪くする.ブルーカットを勧める理屈である.
ただし,実際に昼夜のリズムを狂わせるのは波長よりも明かるさの方だとして,Purkinje現象下の薄明視状態も指摘されている.これらにより夜間のディスプレイには黒基調のダークモードが意味があるとされる.
🗸時計遺伝子 : 交感神経を介して眼圧日内変動を支配している可能性がある.
●皮質拡大機能 cortical magnification :
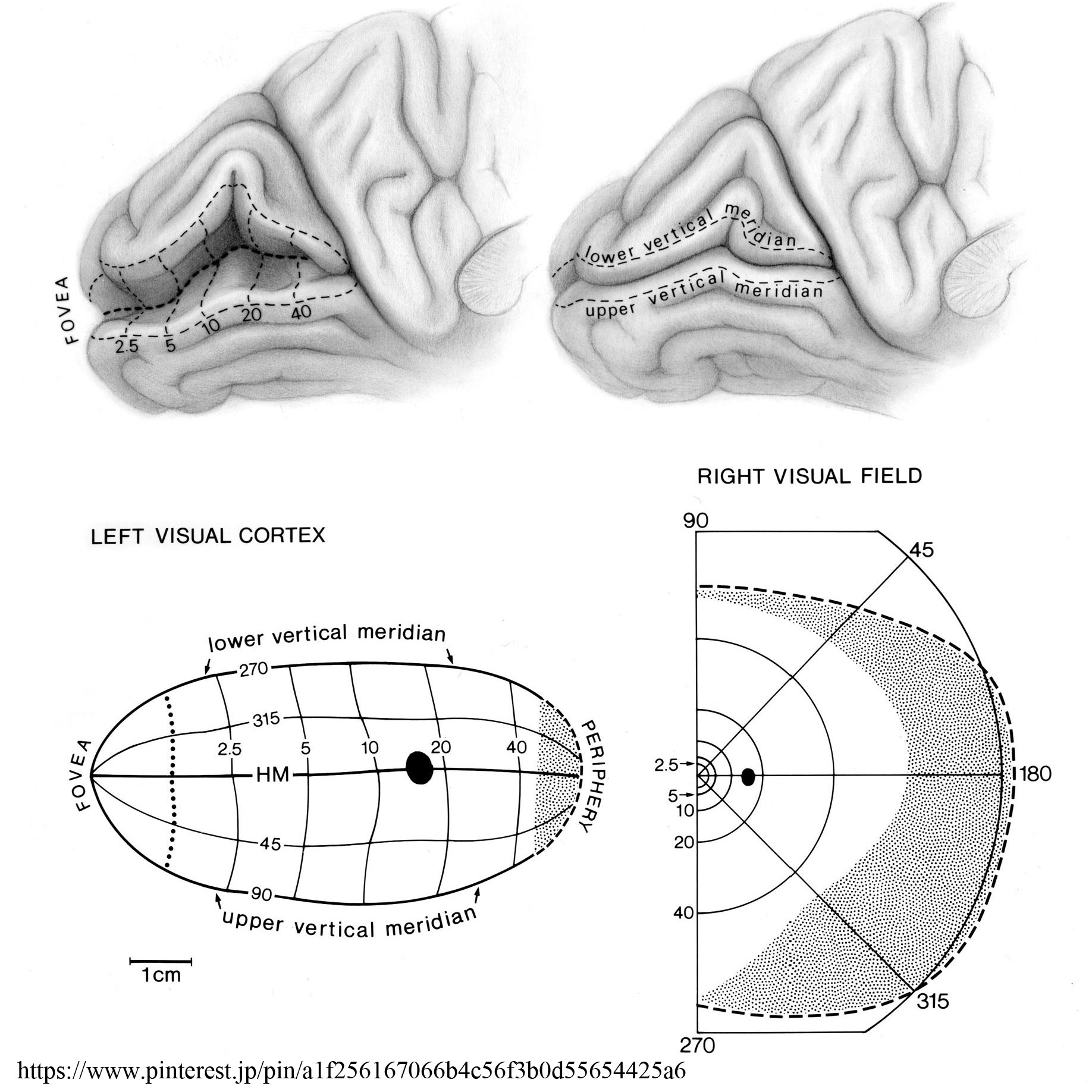
ヒトの視覚系は,中心の狭い範囲を詳細に見ることができるように発達している.黄斑部の神経細胞数はきわめて多く,網膜周辺の神経細胞は少ない.大脳後頭葉の一次視覚野でも後頭極付近にある黄斑部領域をつかさどる神経細胞はきわめて多く,視野の中心2.5度で一次視覚野の30%前後を占め,5度で40%,10度で60%を占めている.視野の40度より周辺では,一次視覚野の幅にして僅か12.5%の範囲に投射されているに過ぎない.
【引用:榛原総合病院学術雑誌 14巻 1号 2019年】
{kind=link}
{kind=link}
{kind=link}