| 一次視覚野 と 高次中枢 |
| primary visual cortex・higher visual cortex |
視覚とは,光情報による視細胞の活動パターンから生体にとって意味のある情報を取り出す機能である.外側膝状体(背側核)から送られた視覚情報は,方位と空間周波数に反応する第一次視覚野・第二次視覚野(低次視覚野)で形状解析・処理され,次いで視覚前野(高次視覚野)に送られる.
また左右の両眼機能の成立には,視覚の知覚系と眼球の運動系の協調運動が必須となる.
外側膝状体ニューロンの軸索は,有線領(線条野・Brodmann17野・V1野)に終わる.
|
|
|
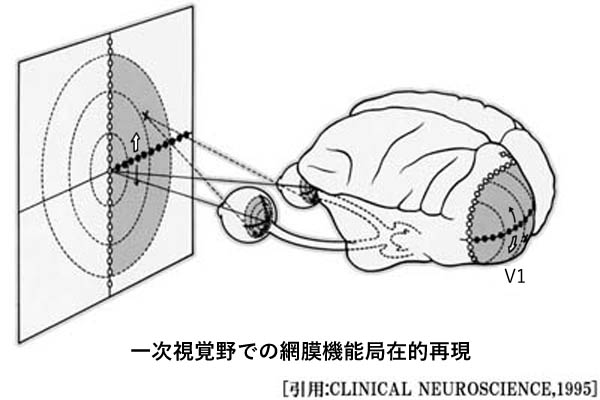
一次視覚野とはV1野を指し,対応する視野空間情報を処理する.それぞれの部位は,分割された情報を同時に処理する単位(モジュール module)の集合である.機能モジュールセットは縦に配置され,各々視野の中でおよそ1°ずつの範囲を担当するという.
機能というのは,担当する部位での分割情報のうちで特定の属性に反応する,という意味である.すなわち,かたち,幅,輪郭の傾き方向,運動方向,色などで,特定の条件にだけ反応する.このことで,モジュールは視覚刺激の特徴検出を担当していると考えられている(ただし,景色がそのままアナログになっている訳ではない).
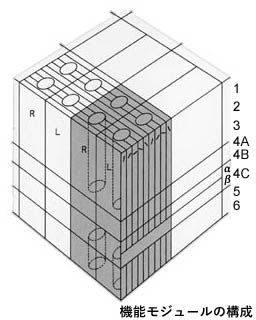
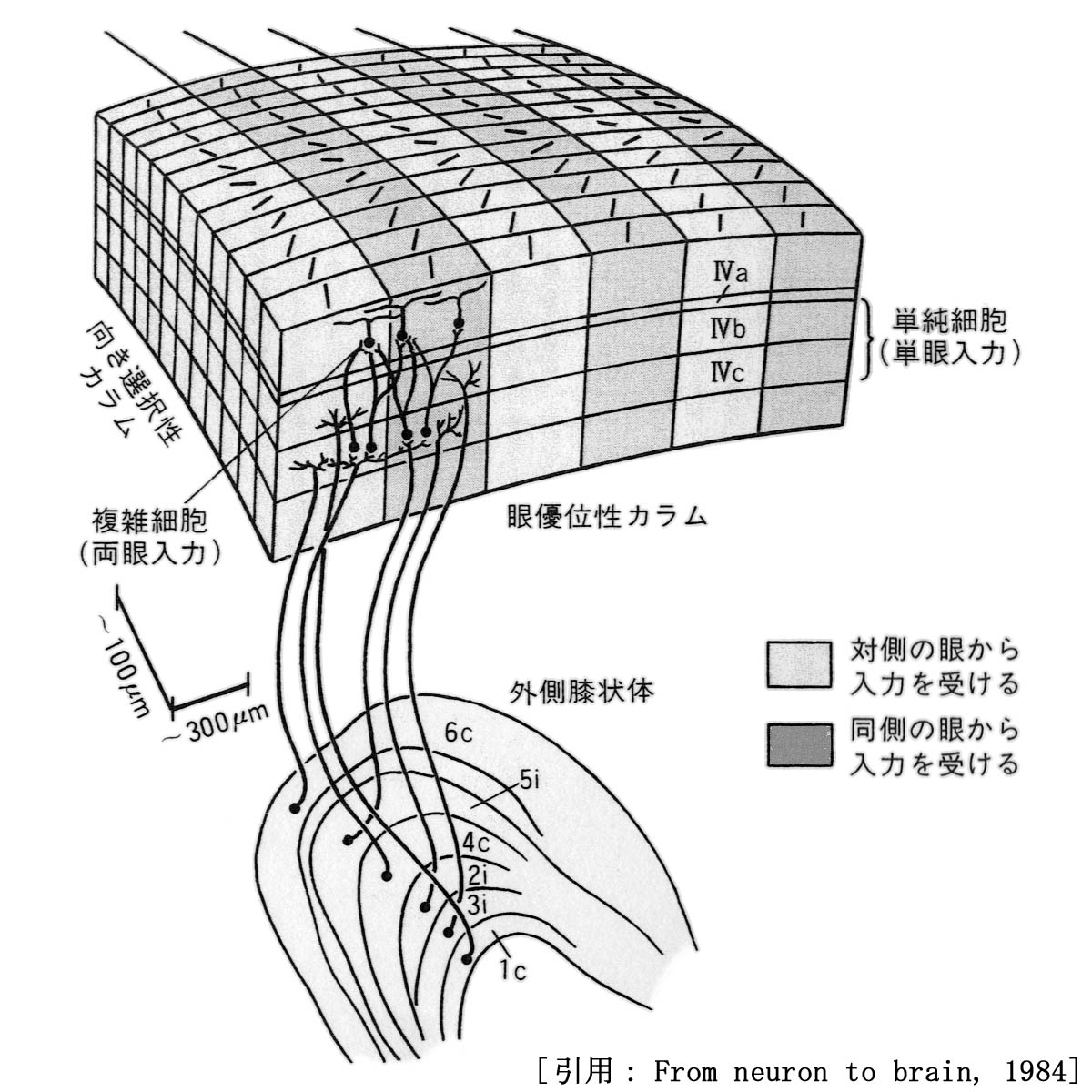
6層で構成される垂直経路.
膝状体からの情報は中層(4C層)で受け取られ,形態認識情報を2・3層の細胞で,眼球運動制御情報を4B層の細胞で抽出する.
各モジュール中のニューロンは視野中の非常に狭いある範囲に含まれる様々な特徴の解析を行なう.
性質の共通したニューロングループは垂直に配列し,機能的円柱(functional column)を形成する.
一定の方向(傾き)に反応するニューロンは,同じ高さに連続している.隣あわせのニューロンは方向の傾き検出が(10°ずつ)連続的にずれ,皮質上の約1mmで全方位180°がカバーされるという.この集合で全視野に対応する.ひとつの方位円柱にスリット・断端・角度に反応する細胞が含まれている.
方位円柱は眼優位円柱と直交する.
コラムには入力信号が強く発現する側,すなわち右信号を受けるニューロンと左信号のそれとが互い違いに配列する.右・左眼優位円柱である.
コラム構造の中で色に反応する部分.線分の方向を検出することはできない.
各々のブロッブは一種類の反対色情報を処理するらしい(赤‐緑,または青‐黄).
17〜19野のニューロンの性質は神経節細胞と大きく違い,特定の図形刺激にのみ反応する回路網になっている(線分方向検出器).
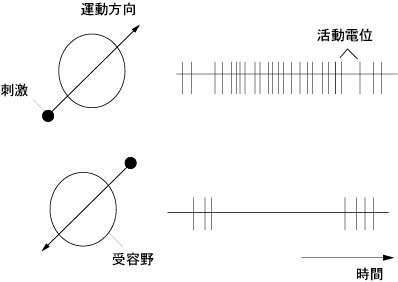 反応特異性から視覚野細胞は三型ある.
反応特異性から視覚野細胞は三型ある.
❶単純型細胞 (simple cell)
中心部に細いスリット状(長方形)のon反応の受容野(on帯)がある.傾き,つまり受容野の長軸方向(角度)を検出する鋭い方位選択性(orientation selectivity)がある.明暗の境を見ているらしい.
❷複雑型細胞 (complex cell)
明確なon帯off帯を持たない.受容野を横切って動く,スリット刺激に応答.幅や運動方向を検出する方向選択性(direction selectivity)をもつ.
❸超複雑型細胞 (hypercomplex cell)
終端抑制の部分を持つ → 断端,長さを検出している.固視微動 に関わると言われる.
以上より視覚情報には階層形式(hierarchy)すなわち,単純型 → 複雑型 → 超複雑型がある.おそらく,画像の輪郭となる明暗を検出し図形の特徴を抽出している(feature detector 特徴抽出細胞).
第一次視覚野は視覚に必要な部位であるが,図形の特徴のほか,色,三次元空間(視野)の中の位置,運動などの特徴抽出によって,単に視野の小領域で何が起きているかを見ているだけといえる.
物体の知覚や視野の全体を認知するには,第一次視覚野の各モジュールで得られたこま切れな情報を統合する必要がある.この過程は視覚連合野で行なわれる.
両眼の像は視差 parallaxのため一致しない.この不一致が perspective すなわち奥ゆき情報となる.
静止画での位置のズレと動画での動きのズレを利用し,両眼から送られてくる二次元情報から三次元情報を抽出する.
大脳皮質視覚野の細胞には両眼から入力を受け,視差に対し異なった反応を示す細胞が存在する.
これらの奥ゆき細胞には,
①対象物とほぼ同じ距離にあるものに反応(等距離細胞),
②固視点より遠くにあるものに反応(遠位細胞),
③固視点より近くにあるものに反応(近位細胞),
があり遠近情報を処理している.
錐体視細胞は波長感受(分光感度)特性の違う三種類があり,Long wave,Middle wave,Short wave ― 長波長・中波長・短波長の帯域感度に応じた光受容信号を発し,信号強度の比で色を識別する情報を得る.600nm
の波長の光が網膜に当たったとすると,おのおのの錐体には光量子吸収に応じ例えば,R=0.7/G=0.4/B=0.2 のような強さの電気信号が発生する.同様に500nm の波長ではR=0.4/G=0.5/B=0.3 のような信号強度になる.この時点で“波長”情報は消え,三錐体にそれぞれ吸収された光量子の数(応答比)に色情報が置き換わることになる.これが “単一変数の原理” principle of univariance である.
各視細胞の感度特性は大きく重複するが,差信号によって限局した帯域情報が得られ,信号圧縮・効率化に有効である.双極細胞以降では異なる種類の錐体の信号の差分(L-MやS-(L+M))をとることにより色の信号が作り出され,この信号は外側膝状体小細胞系経路で伝えられる.一方,信号を加算(主に L+M)することで輝度(明るさ)の信号が作り出され,この信号は外側膝状体大細胞系経路で伝えられる.色信号は外側膝状体の顆粒細胞層(koniocellular layer)でも中継されていると考えられている.
【第九章】 外側膝状体
要は, L 錐体,M 錐体,S 錐体が,それぞれ赤・緑・青(単色光)を認識する,と考えると大きく 誤解 する.
視覚経路のなかに,お互いの反対色のスペクトルに応答するニューロン(反対色細胞)が存在するほか,色情報と形情報とはLGBのレベルよりも中枢側で分離されるらしい.
色覚関連中枢(V4 野)は後頭葉底部の紡錘状回(と舌状回)に存在する.
この部位は,実際の視野では下半視野領域に相当し,両側下半盲を呈する疾患での色覚異常が報告されている.
色情報は錐体細胞で符号化され,圧縮された伝送信号をデコードすることで情報認知していると言えそうである.
●色の恒常性
物体の色は,表面で反射される光の波長分布で決まる.
実際の光は,自然界でも晴天や曇天で違うし,ビルの谷間と海岸でも違う.さらに人工光もあって,これらの光源の差は明らかであろう.しかし我々は,照明光が変わっても物体の色の見えかたはあまり変わらない(ように感じる).例えば,電灯下でも蛍光灯下でも同じような色に見える.これが色の恒常性で,不良設定問題処理の一つである.本来の照明光である白色(=太陽光)を基準として,照明光がもつ配光(分光)特性のズレ(偏り)を脳が修正するらしい.
(カメラのフィルムやCCDなど人工の感光モジュールでは,照明光の特性がそのまま現われる.)
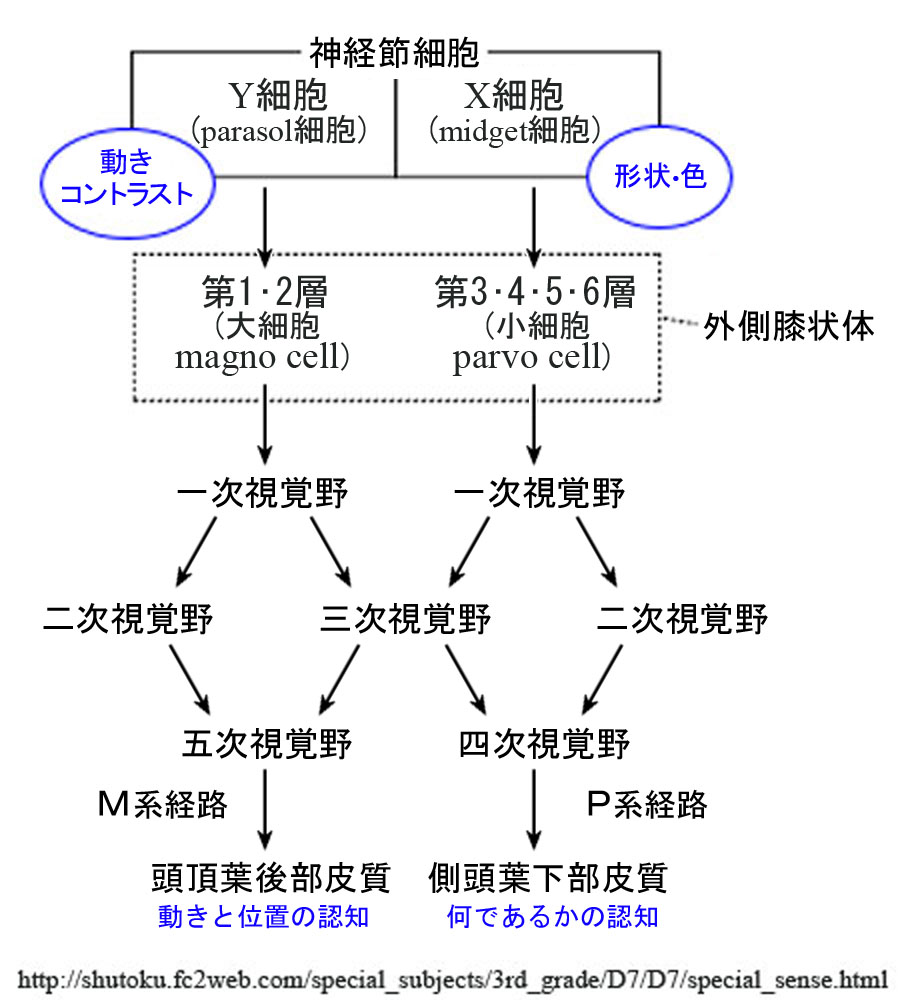
視覚野は機能別に,第一次視覚野(V1),第二次視覚野(V2),第三次視覚野(V3),第四次視覚野(V4),第五次視覚野(V5,MT)に分けられる.
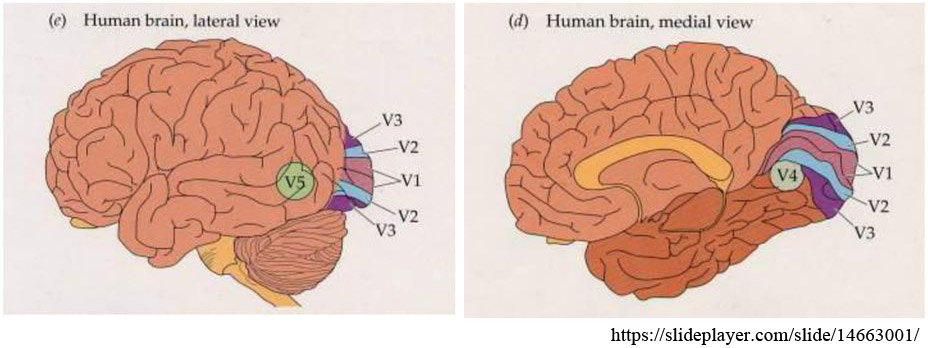
実空間の「モノ」の存在とらえる視覚情報は,「色や形」・「動き」・「奥行き」の三つで処理されると考えられている.ここではさらに単純化して「どこ」に「なに」があるかというように解釈する.「どこ」というのは位置や物体の動きに関する空間情報であり,「なに」というのは物体の形状や色に関する形態情報である.
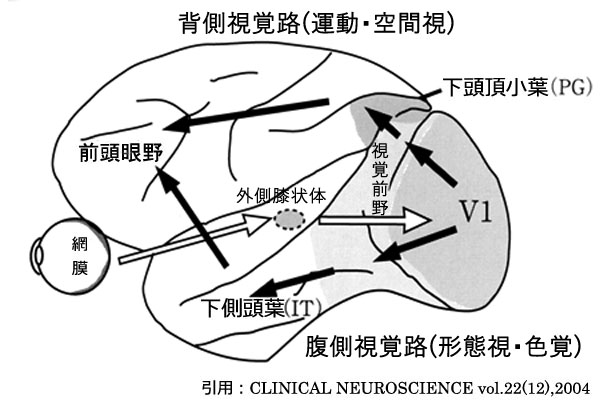
V1に入力した視覚情報は,視覚前野を経て後部頭頂葉皮質や下部側頭葉皮質に伝えられる.すなわち「どこ経路」はV2・V3・V5から頭頂連合野へ出力され,「なに経路」はV2・V3・V4から側頭連合野へ出力される.
①大細胞(M)系チャンネル ~ 位置・動き(高速処理,即ち動態覚 motion vision)
網膜Pα(Y型)神経節細胞→外側膝状体M細胞(1,2層)→V1野4Cα層→4B層
→V2→V3→V5(MT)へ(立体視・運動視の情報処理
②小細胞(P)系チャンネル ~ 色・形(精密処理,即ち形態覚 form vision)
網膜Pβ(X型)神経節細胞→外側膝状体P細胞(3,4,5,6層)→V1野4Cβ層→2・3層
→V2→V4へ(色覚の情報処理
→V3→V4へ(形態視~細かい輪郭の情報処理
まず視野の中で動く物体を見つけるのは“大細胞系”(背側経路),さらに視力を使い詳しく見つめるのが“小細胞系”
両眼視による空間認知の発達・形成は生後 4~5 か月ころ,とのことである.
視神経炎では小細胞系の障害が,緑内障では大細胞系の障害が先行する.
弱視においては小細胞系の不具合がある.
視覚領第2野
立体視:視差(左右眼のずれ)の検出は V1→V2→V3→V5⇒頭頂連合野(背側経路)がメインであるが,V3→V4⇒側頭連合野(腹側経路)にも情報の入力があると考えられている.背側経路では輻湊開散等の運動制御を行い,腹側経路では意識的な奥行き知覚に関連する.

主観的輪郭の検出:現実の画像にない仮想的な(補完)線分に反応を示す細胞がある.
これにより存在しない輪郭(主観的輪郭線 subjective contour)が見える.(右図)
→ エーレンシュタイン Ehrenstein現象という.
スポットの検出:小さなスポットに選択的に反応する細胞がある.機能不明.
それ以降 ・・・
図形の種類や縞の細かさ等,高度な形態認知をしているらしい.
大きい受容野で両眼からの入力を受け,網膜局在性を示すのは後頭連合野までのようである.
さらに19野は,視覚印象の解釈および他の感覚印象との統合に必要である.
 |
 |
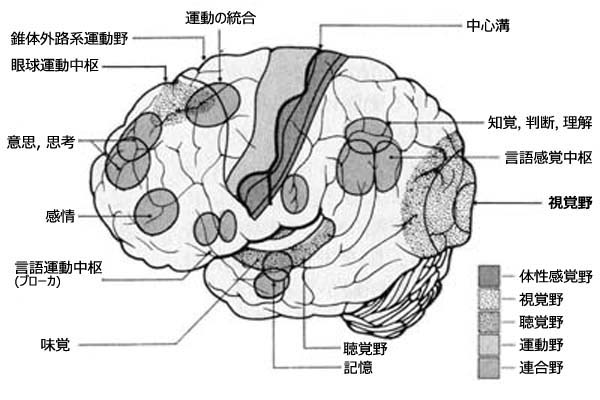
高次領域では,連合野association areaとして情報の統合が行なわれる.前頭連合野,頭頂連合野,側頭連合野,後頭連合野がある.
後頭連合野(視覚連合野 occipital association area:
第一次視覚野の周囲にある皮質 extrastriate cortex(または有線前野 prestriate cortex.18,19野)である.第一次視覚野(V1,Brodmann17野,有線野・線条領)からの入力を受け,視覚情報処理として形態・運動方向の特徴抽出を行なう.17野の前方にあり,視覚前野ともいう.
| V2;第二次視覚野 | 18 野:傍有線領,parastriate cortex |
| V3;第三次視覚野 | 19 野:周有線領・周線条野,peristriate cortex |
| ↓ | (V1〜V3(17〜19)野は“初期視覚野”である) |
| V4;第四次視覚野 (有線前野) | 形(LO;lateral occipital area)と位置(静的な立体視~形態視)・色の識別 → 側頭葉へ(腹側(P系)視覚経路 |
| V5;第五次視覚野 (外有線野) | 視差,対象物の動きや方向(動的な立体視〜動態視)の分析,眼球運動を指令 → 頭頂葉へ(背側(M系)視覚経路 |
*後頭連合野は純粋に視覚情報処理に関係するので,最近は視覚前野として扱い連合野には含めない傾向にある.
*後頭眼野:Brodmann19野を指す.上丘と連絡し,動いている対象物を眼を固定し注視し続ける,追従運動中枢.
20野(視野の地図がある ?)と連動し,近見反応に関わる.視蓋前域で前頭眼野からの下行路と合流する.
側頭連合野(側頭性経路 temporal association area:P系経路
「なに」what情報すなわち対象物の形状認知・形態視に関する視覚情報は,視覚野(V1野) → 視覚前野 → 下側頭回(TE野)に至る.すなわち「形の検出と立体視」である.ここは下側頭野(IT;inferior temporal area)の領域(Brodmann20,21野)で,大脳半球の腹側にあり 腹側路 ventral pathwayである.
二次元情報に加えて奥行き情報も検出するとのことである.これにより運動視情報と他の情報の統合をしている.
また,顔の特徴に反応する顔ニューロンがあるそうだ(顔や全身の動きに反応する,ということで次のMST野からの情報入力がある.また顔ニューロンにより,シミとかカゲがヒトの顔のように認識され易い,との理由にもなっている).
顔認知のほか,記憶・音声言語・聴覚などへの関与.
・側頭経路は女性脳と表現され,例えば色彩認識のような状況が挙げられる.(・・・可否はともかく こんな説明のテキストがあったので紹介)
頭頂連合野(頭頂性経路 parietal association area:M系経路
「どこ」where情報すなわち対象物の位置や移動方向,速度・動態視に関する視覚情報はV5(MT野 MT;middle temporal visual area)から MST野(medial superior temporal visual area)へ入力する.目標の速度・方向と速度情報,すなわち「動きの検出と立体視〜視空間」である.非優位脳半球の下頭頂小葉(IPL;inferior parietal lobule),縁上回 supramarginal gyrus(Brodmann40野)と角回 angular gyrus(39野)に位置し,眼球運動に関与する情報を頭頂葉に送る
背側路 dorsal pathwayである.
頭頂眼野や前頭眼野,前頭前野などと信号のやりとりにより,三次元情報すなわち立体視の中枢となるほか,視野の中の対象物の動きと眼球運動を含む自分自身が動くこと(体性感覚情報)による網膜像全体の動きとを統合し,平行運動,回転運動,拡大・縮小など複雑な視空間情報の処理を行う.
形態覚,聴覚,体性感覚との統合もある.
・頭頂葉は追視している対象物に対する注意に関与する(visual attention).大きく2系統で構成される.
無意識に,位置・形・運動の分析を行う(いかに系;how system)部位は上頭頂小葉(背背側経路),
意識的制御を行う(どこ系;where system)部位は下頭頂小葉(腹背側経路),
・さらに近年では3つに分岐するという.3つの経路(頭頂葉-前頭前野経路、頭頂葉-運動前野経路、頭頂葉-内側側頭葉経路)はそれぞれ,空間的な作業記憶,視覚に基づく動作,記憶に関連したナビゲーションに関わる可能性が指摘されている.
・MT野は,映画・テレビの16~24~30コマ╱秒の静止画のつながりを滑らかな動き(仮現運動)のように認知する場でもある,とのことである.
*頭頂眼野 PEF;parietal eye field:FEFや背外側橋核とともに衝動性追従運動を制御.PEFは特に反射的注視に関与し,スタート信号を上丘や橋に送る.
・頭頂経路は男性脳と表現され,例えば地図を読み解くような状況が挙げられる.
前頭連合野 frontal association area:思考,判断力,社会性,眼球運動の制御などに関連.
*前頭眼野 FEF;frontal eye field:大脳半球外側面の中前頭回の後部の Brodmann8野で,運動前野のすぐ前方にある.上丘や橋と連絡し,視野内の特定の対象物に向ける眼球の随意運動を制御(動きを予測している,とか).水平性眼球運動の皮質中枢で,saccade・smooth pursuit・輻湊に対応する.さらに前頭前野では記憶・判断の処理をおこなう.
その他,投射される部位(視覚領)は多数(30以上とか ?)あり,大脳皮質表面の 1∕4以上に亘っているとのことである.それらの機能は推定・研究段階である.
すなわち,
網膜に入射した光学像は電気信号に変換されて後頭部視覚野(17野)へ伝送される.
さらに大脳の各部に伝送されて,情報処理が進む.
情報伝達の神経回路は空間的に高密度分布で時間応答が遅い経路と,
空間的には粗であるが時間応答の早い経路とがあり,
前者は空間情報を,後者は時間変化情報を主に扱うというように,機能の分化がされている.
また,脳における情報処理過程も脳の左半球では論理・言語を扱い,
右半球では印象・音楽を扱うことが知られている(右利き).
左半球・右半球間は脳梁で連絡されて統一性が保たれる.
画像情報のうち文字・図形は左半球が担当し,
風景・人物などは右半球が担当する.
視覚イメージの処理は特に中心窩部分に集中している.視覚皮質ではニューロンの50%以上が視野の10%しかない中心部の解析にあたっているとされる.このような視覚システムでは,パターン認知システムを視野の意図する部位に向ける方法を持たなければならない.すなわち,明確な視覚を得るには視線がすばやく移動し,常に解像度の高い中心窩で視覚対象を捉えていることが必要である.
これは両眼の正確な協調運動によって可能となる.すなわち,
a)対象を探査・注視する saccade 衝動性眼球運動,
b)対象の移動に合わせて追視するような smooth pursuit 滑動性眼球運動,
c)頭部を動かすなどで手持ちビデオカメラのような「ブレ」を起こさないような vestibular eye movement 前庭動眼反射あるいは optokinetic eye movement 視運動性眼球運動,
がある.
【第十二章】 眼球運動の制御
a)は(注視対象への)視線の移動にかかわる,b)c)は(注視対象への)視線の安定化にかかわる眼球運動といえる.
眼球運動の基本的な皮質中枢は,前頭眼野と後頭眼野である.
視覚反射には,大脳皮質の関与しない皮質下反射と,大脳皮質が関与している皮質性反射がある.
皮質下反射の反射弓は大脳皮質を通らず,眼球の位置の調整と瞳孔の調整をする.求心性線維は視神経,前庭神経,聴神経,脊髄からの上行性線維などを通っている.光の強さにより瞳孔の大きさを調整する対光反射は視蓋前域により,眼球の位置の調整は主に上丘により行われている.
皮質性反射の反射弓は大脳皮質を経過し,眼球の位置を固定する反射と調節反射とがある.眼球の位置の固定には後頭葉が関与している.調節反射はみようとする物体の距離によって,輻湊を変え,瞳孔径を変化させ,調節をして焦点を合わせる反射である.
臨床的に重要なものは対光反応(光反射)と輻湊反応(遠近順応反射)である.
1)対光反射・対光反応
2)近見反射・輻湊反応(近距離輻湊反射 accommodation reflex for near vision 【第十二章】 瞳孔
3)定位チャンネル
視覚イメージから標的の位置と向きを計算する過程は,2つの独立した神経チャンネルが使われているらしい.眼から脳への神経線維の一部が外側膝状核へ向かうものから分かれ上丘に伸びている.上丘を出た神経線維は眼球運動や頭の向き,姿勢の調節などを扱う運動制御システムと結合している.
また,上丘は視覚情報を受容する皮質からも情報を受け取っているらしい.標的を動かすと上丘の細胞は活性化する.視覚・聴覚・触覚から送られた情報は物体を定位するのに役立つ.また,身体の向きを制御するのにも利用されている.
【第九章】 上丘
網膜像から外界を構築する逆光学の過程が,解が不定になる「不良設定問題」である.
われわれが目で見る三次元を網膜では二次元の映像としてとらえている.二次元情報は視神経を介して脳に送られ,脳では二次元の情報を処理・解釈してもとの三次元の世界を推定・認知している.目に飛び込んできた二次元像を三次元空間に置かれた物体として「再現」する過程が,inverse optics「逆光学」である.
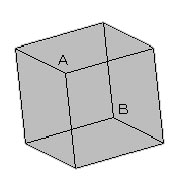
この過程での問題として,三次元の情報を二次元の情報に圧縮・変換すると,必ず情報が失われる.ひとつの二次元図形に対応し得る三次元物体は複数存在し得る.たとえば輪郭線のみで表現された立方体は,どこが手前なのか奥なのか判定しようが無い.図で言えばAが出っ張って見えたりBが出っ張って見えたりと,反転を起こす.
このように,二次元情報をもとにして三次元情報に復元しようとすると,情報不足に起因するよるさまざまな計算上の困難が生じることになる.これが視覚認識の
ill−posed problem 不良設定問題 である.
視覚情報を認知するには、視覚路が完全でなければならない.視放線または第一次視覚野に損傷があると,反対側の視野が見えなくなる.上丘は,手の動きをガイドする部位へ視覚情報を送っているが,意識的に認識をするところには情報を送っていない.しかし,見えなくなっているはずの患者の視野に物体を置き,その物体に手を伸ばすように言うとかなりの正確さでそれを実現できることが知られている.もとは後頭葉損傷者で盲視野内での運動視に対する機能の残存を認めたこと(1917年)で,視覚野障害が強く,連合野皮質が比較的保存された時に観察される.これにより外側膝状体を通らない視覚情報経路の存在が指摘され,その後,膝状体外視覚系として研究されている.
患者は静止しているものは見えなくても,動いている物体を感知できたり動きの方向にもある程度反応できる.これらが
一般には,静的視野と比べて動的視野が広いことをいう.
盲視現象は光は見えないがその方向に目を向けることができる現象である.視蓋部(上丘,副視索核)までは情報が届いている,ことに因るらしい.膝状体外視覚とは,動く対象目標に対してチラツキのような感覚とされる.いずれにしても,意識的な知覚がなくても視覚情報は動作を制御できることを示している.
【 ![]() Riddoch と 盲視 】
Riddoch と 盲視 】
神経回路網の構築は,生涯にわたって絶え間なく改変をうけうるやわらかい構造である.すなわち,回路網に新たな神経要素を加えたり取り除く過程などの再構成に,いろいろな可塑性がある.
これらはおそらく弱視についての診断・治療の可能性に関わることになる.
2024