炎症 とは,生体への傷害因子(刺激・損傷・異物侵入)に対し,内外の異物・病原体の防御や排除,壊死組織の処理,欠損した組織の修復にかかわる一連の生体反応である.これらの過程は,①血管拡張・透過性亢進期,②細胞浸潤期,③細胞増殖期,④組織再構築,と考えられる.本来は生体の合目的的な防御反応であり,自然免疫における重要な要素の一つでもあるが,持続する炎症反応は組織破壊や再構築が過剰に発生することもある.
免疫 とは,主に生体内に侵入した微生物(病原体)を非自己(抗原)として積極的・効率的に排除する生体反応である.非自己には,外来のあらゆる物質のほか自己細胞が変化した腫瘍細胞などを含めて考えることができる.これらの多様性と特異性において,
さらに,❹多様性を生み出すために遺伝子の再構成を行う.
免疫システムは,⑴自然免疫,⑵適応免疫(獲得免疫),がある.両者の連携が重要である.
![]()
免疫は自己(self)と非自己(notⲻself=foreign)を識別し,(有害,と認識した)非自己を区別・排除すること,すなわち自己防衛機構である.関与する細胞は白血球グループとして包括する.
免疫反応に基づく生体に対する全身的または局所的な障害が アレルギー allergy で,多くが本来ならば無害な非自己に対する過剰(または,誤作動)反応である.自己成分に対する区別・識別の狂いが 自己免疫疾患
生物学的な自己主張は,細胞表面の
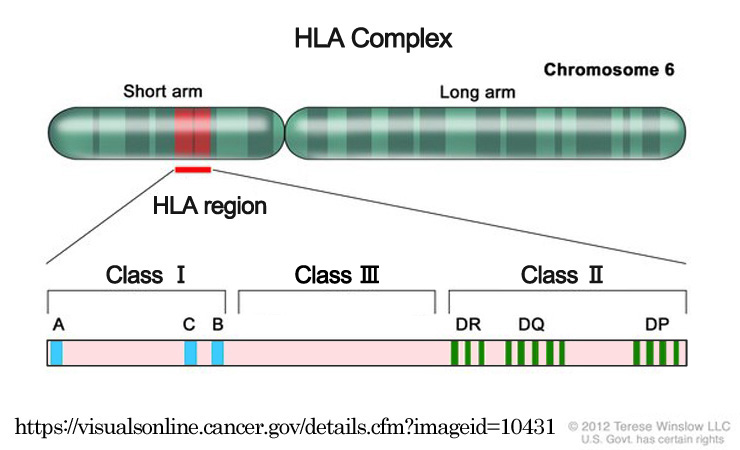
HLA に依る.
免疫系はこの 目印 をみて自己 self か非自己 notⲻself かを識別・区別する.
◇感染の成立 infection:
健常者において排除されずに定着・増殖(寄生体 parasite),あるいは,宿主 hostに固有の症状・症候を作り(生体防御反応),病害がもたらされた(いわゆる,発症)状態が “感染症”である.
◇自然免疫の回避:
細胞内寄生体(マクロファージに感染してその内部で増殖する.Salmonella属),
プロテクタで貪食をまぬがれる(Mycobacterium tuberculosis),
レセプタのふりをして貪食を抑制,あるいは宿主細胞をまねて異物認識をまぬがれる(ある種の腸内細菌叢 ― 共生細菌),
貪食能を抑制(Staphylococcus aureus),
貪食細胞を直接傷害(M.tuberculosis,Streptococcus pyogenes,Bacillus anthracis)
◇バイオフィルム:免疫細胞の攻撃を防ぐ構造.
◇遺伝的素因:HLA
◇環境的要因:日光暴露,粉塵暴露,薬剤,加齢,ウイルス感染,喫煙,性ホルモン関連
◇自然免疫に関連する遺伝子異常
◇原因となる病原体や自己抗体,抗原特異的T細胞は検出されない.
免疫応答を誘発した抗原は,非自己と認識されたことになる.
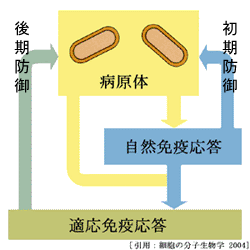
生体防御態勢は大きく二つある.
❶マクロファージ・樹状細胞・好中球・ナチュラルキラー(NK)細胞・自然リンパ球(ILC)等が担う自然免疫系と,
❷リンパ球(T細胞サブセット・B細胞)による獲得免疫(適応免疫)系,である.
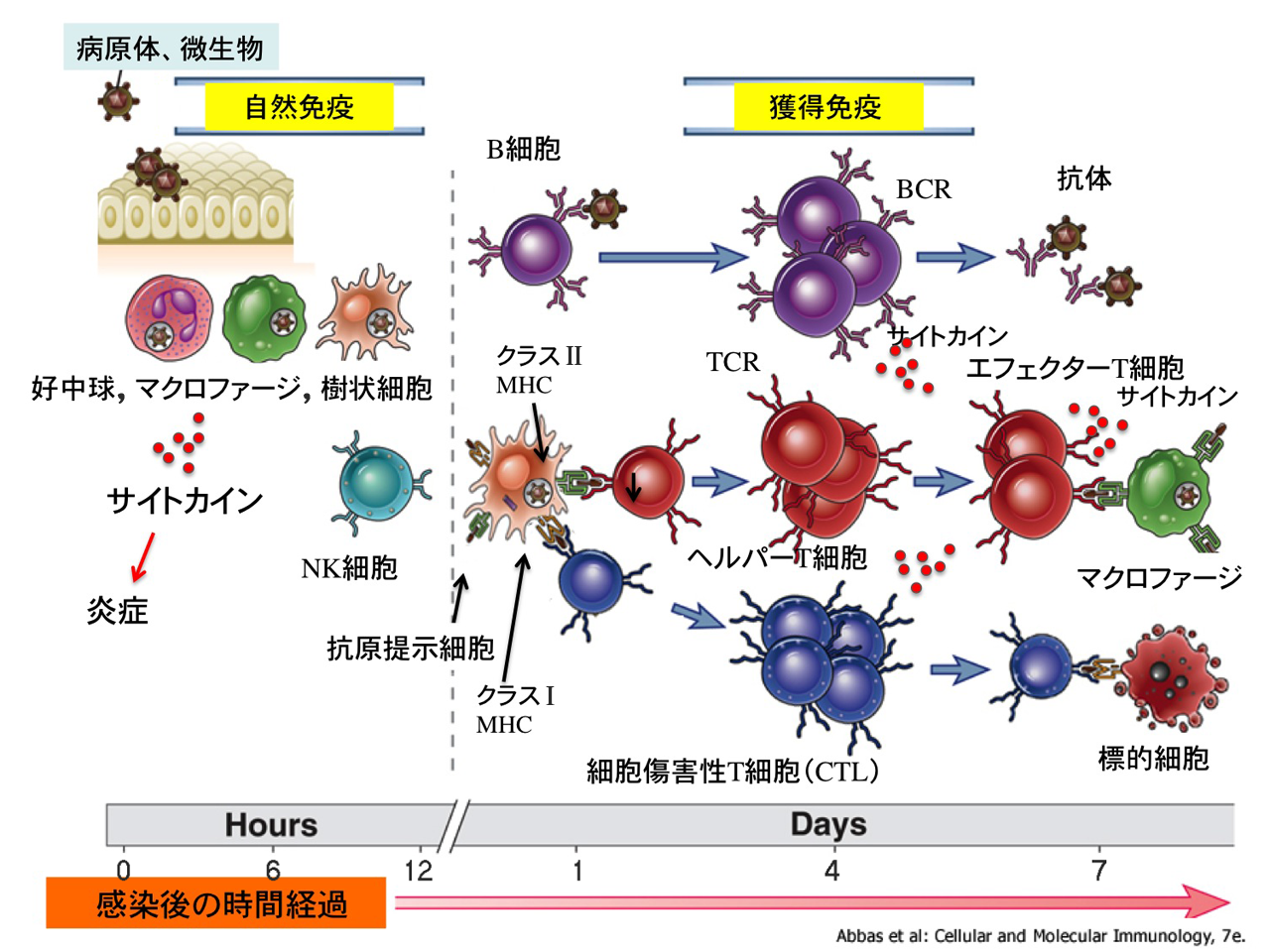
ⅰ)自然免疫応答 innate immunity
抗原非特異的に,外からの異物を宿主にはない型(非自己)と認識し,微生物(感染病原体)に対する初期防御を行う物理的および化学的障壁である.感染(侵入)後,数時間で発動される.抗原特異的な受容体を発現せずに,主に食作用と炎症反応により排除する.獲得免疫への橋渡し(抗原提示)にも重要なプロセスである.
ⓐ一次防衛線:侵入部位(皮膚・粘膜表面)では,上皮細胞(皮膚・気道・消化管 ・・・)のバリア(密着結合構造やデフェンシン・クリプトシジンなどの抗微生物物質の分泌)や上皮内リンパ球が作用する.皮膚や粘膜表面にいる共生微生物・常在菌も病原体の侵入を阻止している.
眼科領域では角結膜上皮や涙液に殺菌物質リゾチームを含み,防御前線となっている.また,好中球が輪部の血管を通って角膜に達し,食作用を発揮する.
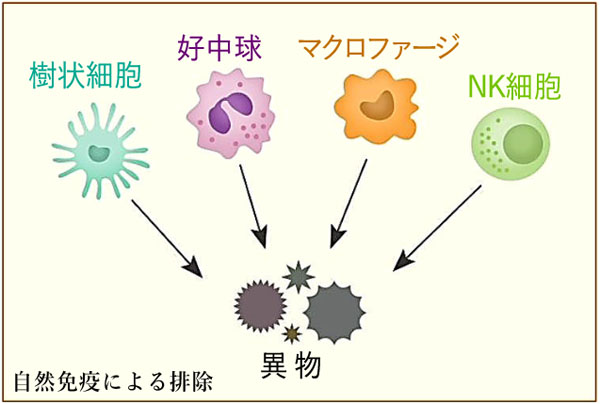
ⓑ二次防衛線:急性炎症反応と食作用がある.関与因子は,❶貪食細胞(好中球・マクロファージ・樹状細胞)やパターン認識受容体 pattern recognition receptors
病原体センサーとしてのPRRは Toll様受容体(TLR)で代表され,主にマクロファージや樹状細胞などが微生物の持つ共通した構成ペプチドなど分子構造(病原体関連分子パターン pathogenⲻ
PAMPs は,病原体に固有に存在しそれらの生育に必須の成分であるが,我々の身体には存在しない構成成分を指す.PRRs による PAMPs認識が免疫発動の起点となっている.
DAMPs は傷害された細胞や組織から放出される危険シグナルで,病原体の関与しない無菌性の炎症反応(sterile inflammation)を惹起する.
これらにより,①特異性の低いブロック(程度は低いが,特異的な病原体の認識機構も存在),②早い応答,③獲得免疫へのリンク,④遺伝子レベルで制御され常に機能している,などが特徴になる.感染を繰り返しても免疫学的な記憶は成立せず抵抗力が高まることもないが,適応免疫の活性化には先行する自然免疫の活性化が必須である(抗原提示 ・・・ 自然免疫から獲得免疫へのリンクが TLRや 樹状細胞 とのことである).
自然免疫は数時間~数日間が勝負であるのに対し,適応免疫応答の立ち上がりには数日間~数週間を要する.
⯎ パターン認識受容体:
【 ✋ PRRs 】
Tollⲻlike receptor(TLR)ファミリー,
retinoic acidⲻinducible geneⲻI(RIGⲻI)様受容体,
インフラマソーム等が知られており,それぞれ異なる病原体成分を認識する.
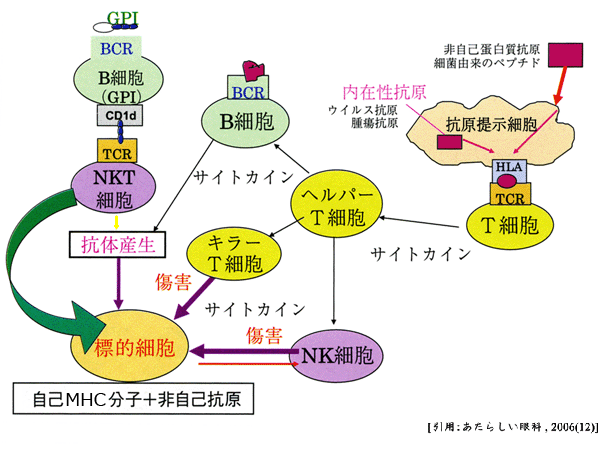
⯎ 自然免疫から獲得免疫へ繋ぐ,もう一つのシステムが NKT細胞 となる. NK細胞とT細胞の両方のマーカーを持ち,抗原受容体はCD1d分子に結合する糖脂質を認識することで活性化する. NKT細胞は多量のサイトカインを産生できることから,NK細胞,T細胞およびB細胞を活性化させる賦活化(アジュバント)機能が生体内での役割であると考えられている.
⯎ 自然免疫系は「感染性疾患」のみならず,気管支喘息をはじめとする様々なアレルギー性疾患,さらには生活習慣病など,多くの「非感染性疾患」においても重要な役割を果たしていることが明らかになりつつある.
通常,自然免疫の活性化は,Th1,Th17 細胞へと分化する方向,すなわちアレルギーを抑制する方向に作用する.時に Th2 細胞へと分化誘導させる場合がある.この Th2 分化誘導が過剰になった状態がアレルギーである.
⯎ 炎症性サイトカイン:ILⲻ1,ILⲻ6,TNFⲻαなど
ⅱ)適応免疫応答 adaptive immunity
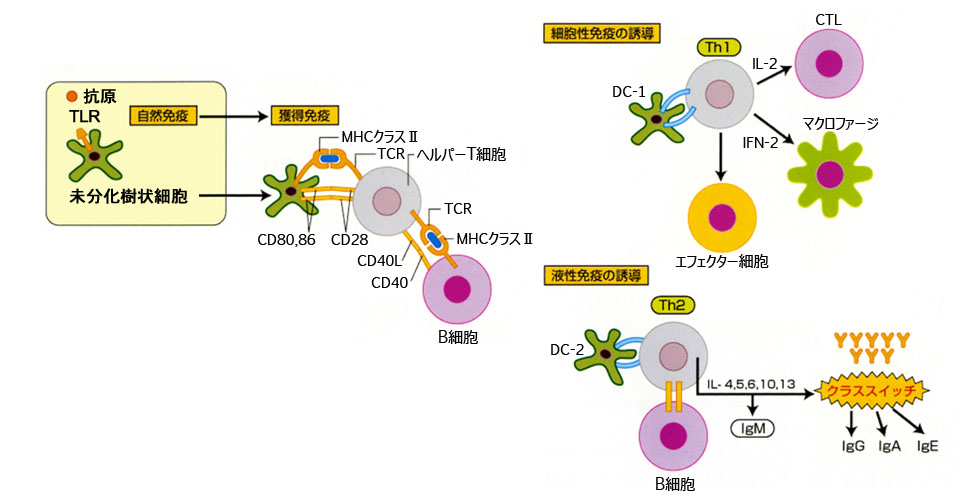
抗原を認識した樹状細胞は最寄りの(所属)リンパ節でナイーブ(naïve;幼弱)T細胞に抗原提示をし,エフェクターT細胞として獲得免疫が始動する(一次免疫反応).
ⓒ三次防衛線:適応免疫応答は細胞性免疫(キラーT細胞主体:感染細胞を傷害)と液性免疫(B細胞主体:形質細胞として抗体産生)の2種類の型があり,侵入した特定の病原体とそれが作る毒性分子を破壊排除する抗原特異的な働きである.マクロファージがおもに末梢リンパ器官を活性化させ,リンパ球による免疫反応が発生する.これらにより “免疫の多様性”を獲得している.
関与因子は,①異物認識(抗原提示),②リンパ球とその産物,である.すなわち,抗原によって免疫系が発動するが,数日から数週間を要する.抗原提示細胞 antigen presenting cell(APC)は,樹状細胞・マクロファージである.抗体,主要組織適合性複合体(major histocompatibility complex;MHC)分子,T細胞抗原レセプタ(TCR)の3種類によって抗原を認識する.
抗原情報を受けたT細胞はサイトカインを産生し,これを受けてB細胞は 形質細胞 に分化して特異的に抗体を産生し,病原体に対抗する(二次免疫反応).
感染を繰り返すか抗原の投与(能動免疫;いわゆる予防接種)により,あるいは抗体の投与(受動免疫;いわゆる母体抗体)により特定の抵抗力が高まる. 獲得免疫を担当する T 細胞や B 細胞は,遺伝子再構成により生み出された抗原受容体を介して抗原認識を行い,その多様性や記憶が大きな特徴である.
適応免疫応答の主役細胞は リンパ球 である.
リンパ球は骨髄のリンパ系幹細胞から2通りに分化し,一次リンパ器官(胸腺 thymus と骨髄 bone marrow)で成熟し,リンパ腺(二次リンパ器官)で休眠し,起こされるとリンパ管や血管を通って体内を循環し,免疫現象で最も重要な役割を果たす.
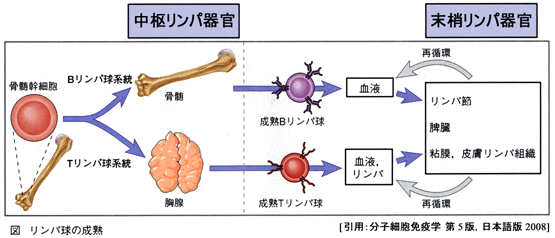
| 骨髄幹細胞 (前駆細胞) |
→ | 胸腺:T細胞 |
| 骨髄:B細胞 |
リンパ球は,抗原によって活性化する.
体内に侵入した外来異物を自分自身と区別して識別する,リンパ球の表面に発現している分子が抗原受容体である.
BCR とは抗体のことである.これにより B細胞は特異的に異物認識をする.
TCR は,抗原提示細胞または標的細胞の表面で結合した MHCと抗原ペプチド(タンパクの断片)の複合体を認識する.断片とは抗原タンパクが細胞内で分解(プロセッシング)されたもの.
単球(マクロファージ)・組織内の樹状細胞(dendritic cell)・皮膚Langerhans細胞・活性化B細胞 などはTh細胞に抗原を提示する.マクロファージや樹状細胞は非特異的に異物認識をする.細胞内で処理した抗原ペプチド(十数個のアミノ酸からなる)を細胞表面でMHC抗原と共にT細胞に提示する.抗原決定基がエピトープ
貪食:
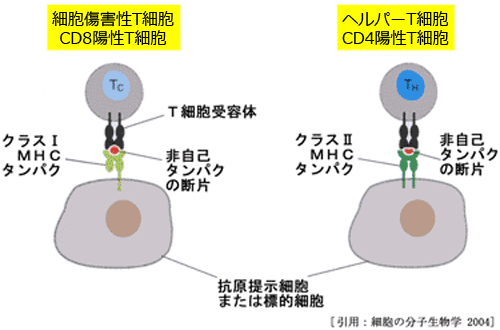
細菌などは単球・マクロファージにより貪食され,細胞内でリソソームにより抗原ペプチドに分解(processing)される.細胞表面のMHCクラスII分子は,抗原ペプチドを挟む形でCD4陽性のTh細胞のTCR・CD3複合体に抗原を提示する.TCRの抗原認識のシグナルは,CD3分子を介してT細胞内に伝達される(シグナル1).Th細胞がMHCクラスII分子に結合した抗原を認識する際には,CD4抗原が補助的に働く.
ウイルスなどの(腫瘍細胞を含む)抗原は,抗原ペプチド(抗原フラグメント)の形で単球・マクロファージのMHCクラスI分子と共にCD8陽性のTc細胞に提示される.CD8陽性T細胞は,クラスI自己抗原を認識し攻撃を避ける.
好中球は貪食・消化後,数時間で死滅し膿となる.
エフェクターTとサブセット:(細胞性免疫 cellular immunity;T細胞免疫)
T細胞抗原受容体(TCR)α・β鎖が,抗原提示細胞表面でMHCにより提示される複合体ペプチドを認識する.
◙ Th細胞/ヘルパーT・・・ HLAクラスII分子と結合した抗原ペプチドを認識し,B細胞の増殖・分化を促す.細胞外微生物に対する宿主の防御に機能する.存在するサイトカインなどの細胞外環境によりTh1,Th2,TFH,Th17,Treg細胞(相互転換もある)などのそれぞれ特異的な機能を持ったCD4陽性T細胞へと分化する. CD4陽性T細胞は,外来抗原特異的獲得免疫応答に必須のリンパ球である.
◙ Tc細胞/キラーT(細胞傷害性;cytotoxic またはキラー;killer)・・・ HLAクラスI分子と結合した抗原ペプチドを認識し,細胞傷害性(ウイルス感染細胞,腫瘍細胞,非自己細胞)に作用する.サイトカインを浴びると増殖し,かつ感染細胞を破壊する力がアップする.CD8陽性T細胞
◙ T細胞活性化(IL-2産生)には,T細胞表面のインテグリン(補助受容体:CD28やCTLA-4)が単球・マクロファージ表面に発現されるインテグリンリガンド(補助刺激分子,B7-1)で刺激され生じる抗原非特異的な シグナル2(共刺激シグナル:補助刺激シグナル)が必要である. これにより,Th細胞の細胞質で IL-2 mRNAの転写が起こり IL-2 が生産され,T細胞が活性化される(エフェクターT).
Bリンパ球:(液性免疫 humoral immunity;B細胞免疫)
B細胞では細胞表面の免疫グロブリン(IgM)が抗原特異的受容体(BCR)として働いて,抗原を認識する.B細胞レセプター複合体によりシグナル伝達が開始され,形質細胞(エフェクターB細胞)とメモリーBに分化し,Th2リンパ球の存在下で抗体を産生する.
B細胞は基本的にIgMが発現しているがTh2が作用すると形質細胞に変身してIgG・IgEほかを産生する.クラススイッチ(クラスチェンジ)という.
免疫の記憶:記憶細胞
異物 → マクロファージが貪食・抗原提示 → ヘルパーT細胞が感知・抗原認識,のプロセスが免疫応答である.通常,『抗原』・『免疫応答』という語は適応免疫を示す.
抗原提示により T細胞 と B細胞が活性化し,それぞれ,細胞性免疫と液性免疫が発動する.
細胞性応答は,抗原情報 → T細胞(キラーT)へ伝達・増殖 → 局所へ移動し,サイトカイン(IFN-γ)などを分泌しその部位のみで反応,抗原を排除する.キラーT細胞は,ウイルス感染細胞のほか結核菌やニューモシスチスを含む真菌に対応する.
液性応答は,抗原情報 → B細胞へ伝達 → 分化した形質細胞(plasma cell)が抗体(免疫グロブリン(immuno-globulin;Ig)を分泌する.抗体は血流に乗って全身で反応する.IgGが菌に結合(オプソニン化)すると貪食能が上がる(オプソニン効果).貪食の残骸が膿となる.B細胞はおおよそ,化膿性炎症をおこす細菌や毒素に対応する.
分泌型IgAを中心とする「粘膜免疫」は粘膜の表面など外界に接する最前線で,病原体などの抗原物質が体内に入ることを防いでいる.この「粘膜免疫」と,体内に入ってしまった抗原物質に対するIgGによる免疫反応とが,液性免疫の二本柱といわれる.
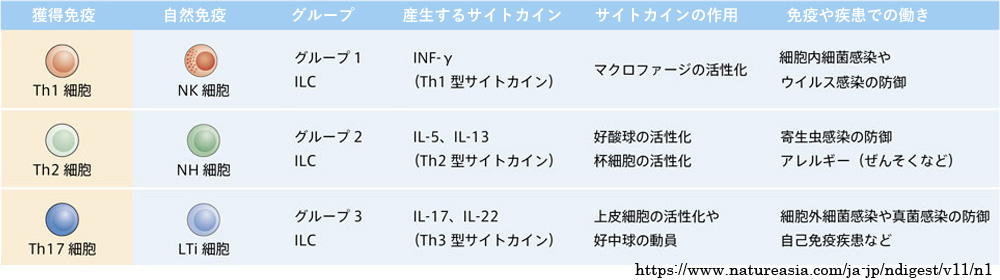
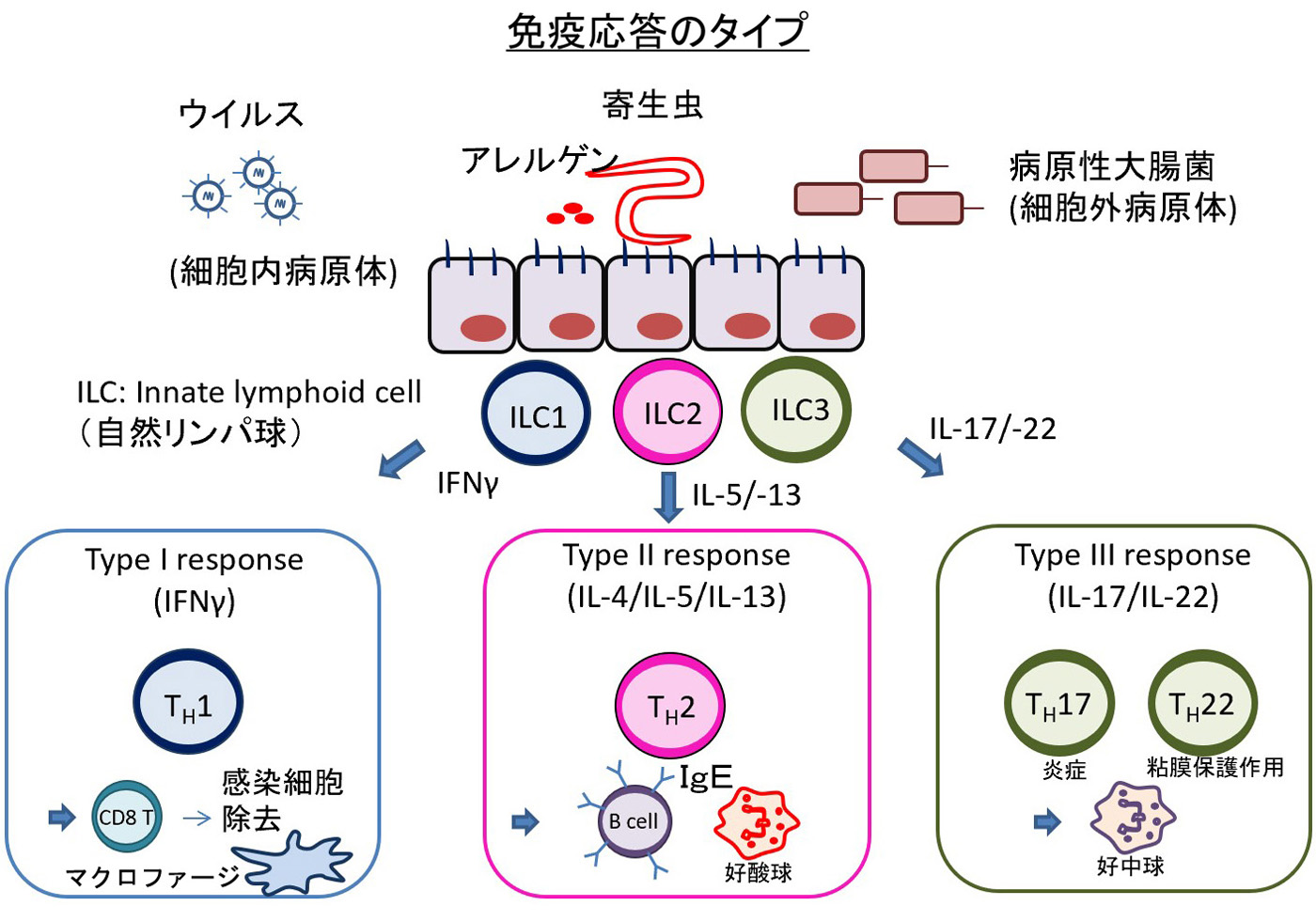
➊1型炎症:IFNⲻγを産生しマクロファージの活性化などを介し,細胞内細菌やウイルスの感染防御を担う. Th1細胞・ILC1細胞が関与する通常の免疫反応.
➋2型炎症:ILⲻ5 や ILⲻ13 を産生し寄生虫の感染防御や Ⅰ型アレルギー反応を担う. 2型炎症応答は寄生虫の感染や,アレルゲン・毒素などの刺激により引き起こされる炎症応答である. 好酸球や杯細胞の活性化を介し,寄生虫(蠕虫)の排除・防御,アレルギー性炎症の発症をきたす. 主体となる2型サイトカイン(ILⲻ4・ILⲻ5・ILⲻ13)の産生は,Th2細胞(獲得免疫),ILC2細胞(自然免疫)による.免疫グロブリンE(IgE)の関与もある.
➌3型炎症:ILⲻ17 や ILⲻ22 を産生し好中球の動員や上皮細胞の活性化・増殖を介し,細胞外細菌や真菌の感染防御を担う.
Th17細胞,ICL3細胞の関与.
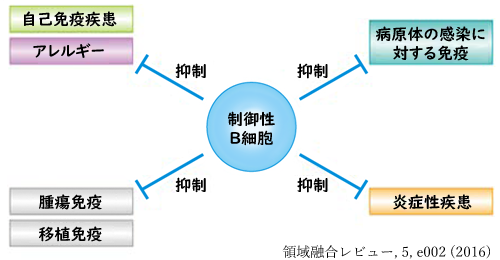
◙ レギュラトリーB:
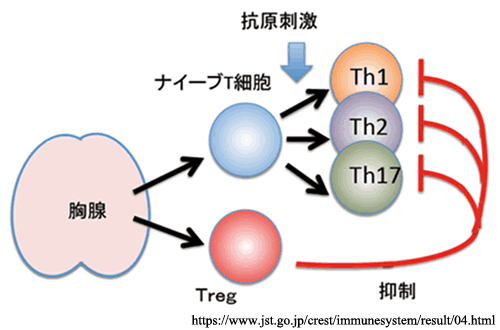
◙ レギュラトリーT:
病原体情報はメモリーされ,長期間持続する.
◙ メモリーB細胞:次回の抗原侵入に備える.抗原感知時には速やかに形質細胞に分化する.
◙ メモリーT細胞:長期間体内を循環し(監視機構),次回の抗原侵入に対する反応を迅速に起こす.
メモリーT細胞は,幹細胞メモリーT細胞(TSCM)・セントラルメモリーT細胞(TCM)・エフェクターメモリーT細胞(TEM)・組織常在型メモリーT細胞(TRM)のサブセットに分類
抗原の除去により免疫応答には自然抑制が働き,免疫系は休止期の状態に戻る.反応の沈静化,すなわちサイトカインが減る(免疫応答の生理的な減衰)とリンパ球は生存刺激がなくなり,炎症反応に関わったリンパ球はアポトーシスに至るが,一部がメモリ細胞として残る (再循環リンパ球:90%以上がT細胞).
加齢とともに蓄積する慢性的な軽度炎症は,感染症とは無関係に臨床的には自覚されないまま,獲得免疫系と自然免疫系に影響を及ぼす.
MHCは,自己を「攻撃対象ではない」と認識する遺伝子群の総称である.ヒトでは,白血球で最初に確認され 『HLA』 と言う(HLA抗原:human leukocyte associated antigen).
主な作用は,自然免疫の制御 / NK細胞の活性化,獲得免疫でのT細胞への抗原提示,である.一般的には,
MHCタンパク は構造と機能によって大きく2種類に分けられる.
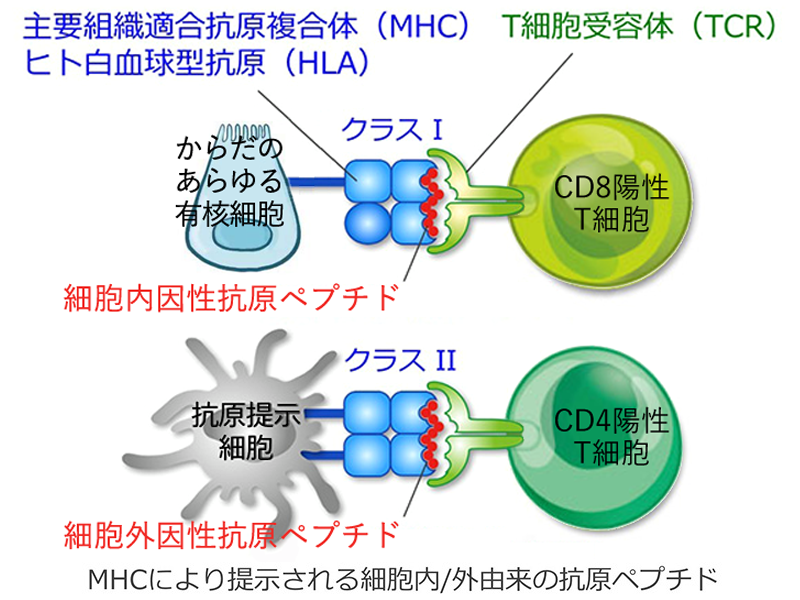
⒈クラスIMHC:HLA-A,-B,-C,-E,-F,-G
全ての細胞表面に発現.血小板表面やT細胞・B細胞を含む(古典的HLAクラスI分子:A,B,C.).特定の細胞や分化段階で発現(非古典的HLAクラスI分子:E,F,G.).
クラスIMHC分子は細胞内在性のペプチド(内因性抗原:自己タンパク,ウイルス抗原や腫瘍抗原など)をCD8陽性T細胞(キラーT)に提示する.クラスⅠMHCタンパクと結合するのは,キラーT細胞に発現した CD8
⒉クラスIIMHC:HLA-D,-DP,-DQ,-DR
樹状細胞(皮膚Langerhans細胞など),単球・マクロファージ,B細胞などの抗原提示細胞に発現(古典的HLAクラスII分子:DR,DQ,DP.非古典的HLAクラスⅡ分子:DM).活性化T細胞(静止期のT細胞には発現していない),胸腺上皮細胞,精子などに発現(非古典的HLAクラスII分子:DO.).
クラスIIMHC分子は細胞外から取り込まれ(貪食し)た可溶性タンパクのペプチド(外来性抗原)をCD4陽性T細胞(ヘルパーT)に提示する.クラスⅡMHCタンパクと結合するのは,ヘルパーT細胞に発現した CD4である.
Th細胞は,キラーT細胞(CD8細胞)などの他の免疫系細胞にシグナルを送る.
| HLA | 遺伝子座 | 抗原 | 抗原提示先 | 機能 |
| クラスⅠ | 第6番染色体短腕部 | 内因性抗原 | 細胞傷害性T細胞 | おもに細胞性免疫 |
| クラスⅡ | 外来性抗原 | ヘルパーT細胞 | おもに液性免疫 |
⒊クラスIIIMHC:補体をコード,自然免疫に関与.
その他:熱ショック蛋白(HSP70),腫瘍壊死因子(TNF),LMP,TAP,・・・・・
◙ ある特定のMHC(HLA)型と相関して罹患率が高い特定の疾患がある.特定のMHC分子の遺伝子型では自己抗原ペプチドを自己反応性T細胞に対して効率よく提示できずに,十分に自己応答性のT細胞の排除が起こらなかった結果であると説明されている.
◙ 表記;抗原からアリルへ 👇
◙ CD(cluster of differentiation: 👇
骨髄から胸腺へと移入してきたT細胞の前駆細胞はそこで分化成熟することになる.
自己の目印は細胞表面のMHCである.免疫系はこのMHCをみて自己 self か非自己 nonself かを識別する.
自 己:MHC+自己のペプチド
非自己:MHC+外来ペプチド
リンパ球の形成過程でMHCに対する反応性リンパ球が消滅するため,自分の組織に対して反応することはない.この性質を,免疫学的自己寛容 immunological self−tolerance または免疫寛容という.
胸腺において,自己MHC分子と全く反応しないT細胞は死滅する.MHCクラスIを認識するT細胞受容体を発現する細胞はキラーT細胞へ,MHCクラスIIを認識するT細胞受容体を発現する細胞はヘルパーT細胞へ分化が誘導される。
B細胞は自己のタンパク質と反応するものはアポトーシスにより死滅し,骨髄外に出ない.
T細胞は樹状細胞上のMHC自己ペプチドと反応するものは胸腺外に出られない(不活化・消去される.
*
アポトーシスの機能不全は,種々の原因により不活化/抑制機構が解除された活性化自己反応性リンパ球の増加を意味し,その結果,自己免疫反応の持続・遷延化をきたして,遂には不可逆的な自己免疫疾患を発症すると考えられる.
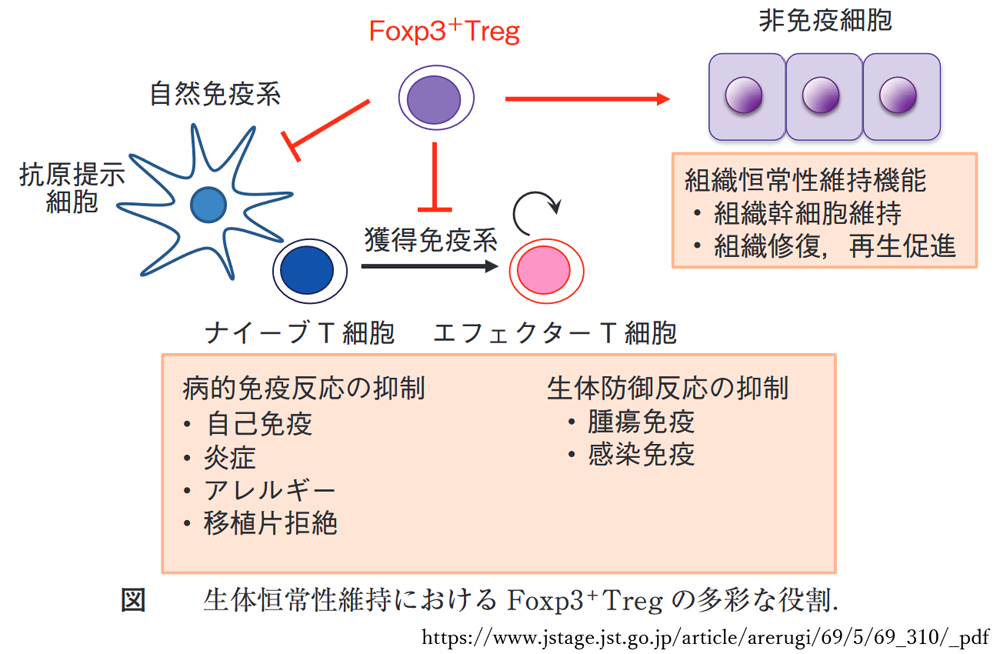
・時折,消去をまぬがれる細胞は,制御性T細胞へと分化する.
制御性T(Treg)細胞による免疫抑制.→ この異常が,免疫寛容の破綻.
無応答・無視.
*発生の過程で免疫系と接触しないまま分化する組織が隔絶抗原である.新参抗原と認識されてしまうと自己免疫疾患を発症する.
欠失.自己反応性T細胞・自己反応性B細胞が消去・排除される.
*Fas(CD95)などによる.Fasの変異があると自己免疫疾患を発症する.
不活化・不応答性.T細胞は,抗原認識のシグナル1のみが伝わりシグナル2(補助シグナル)が伝えられないと活性化されないだけでなく,その抗原に対して応答しない免疫寛容状態になる.補助刺激分子(CD86)は,抗原提示細胞(マクロファージなど)のみが発現している.末梢組織の細胞がMHCクラスII分子に自己抗原を結合して提示(シグナル1)しても,ヘルパーT細胞は補助受容体(CD28)の刺激(シグナル2)がないので活性化されず無反応(anergy)に終わる.これによりヘルパーT細胞は「自己」の成分(自己抗原)には免疫応答(自己免疫)を起こさない.
自己反応性B細胞は反応しないレセプターに変わる(エディティング).寛容状態のヘルパーT細胞ではB細胞は抗体産生が不十分となる.
活性化シグナルを受けられないリンパ球は死滅する.
・能動免疫
・受動免疫
・抗体接種
・自己抗体が発現するようになり,自己免疫疾患の頻度が高くなる.
・マクロファージの機能低下により,腫瘍細胞が消去しきれなくなる.
・T細胞の抗原に対する反応が緩慢になる.
・あらたな抗原に応答できるリンパ球が少なくなる.
・細菌感染に対して産生される補体の量が少なくなる.
・抗体の濃度そのものが著明に低下するわけではないが,抗原と抗体の親和性が低下することが一因で,高齢者における肺炎,インフルエンザ,などの罹患率が上昇している可能性がある.これらの変化により,高齢者でワクチンが効きにくい理由がある程度説明できる可能性もある.
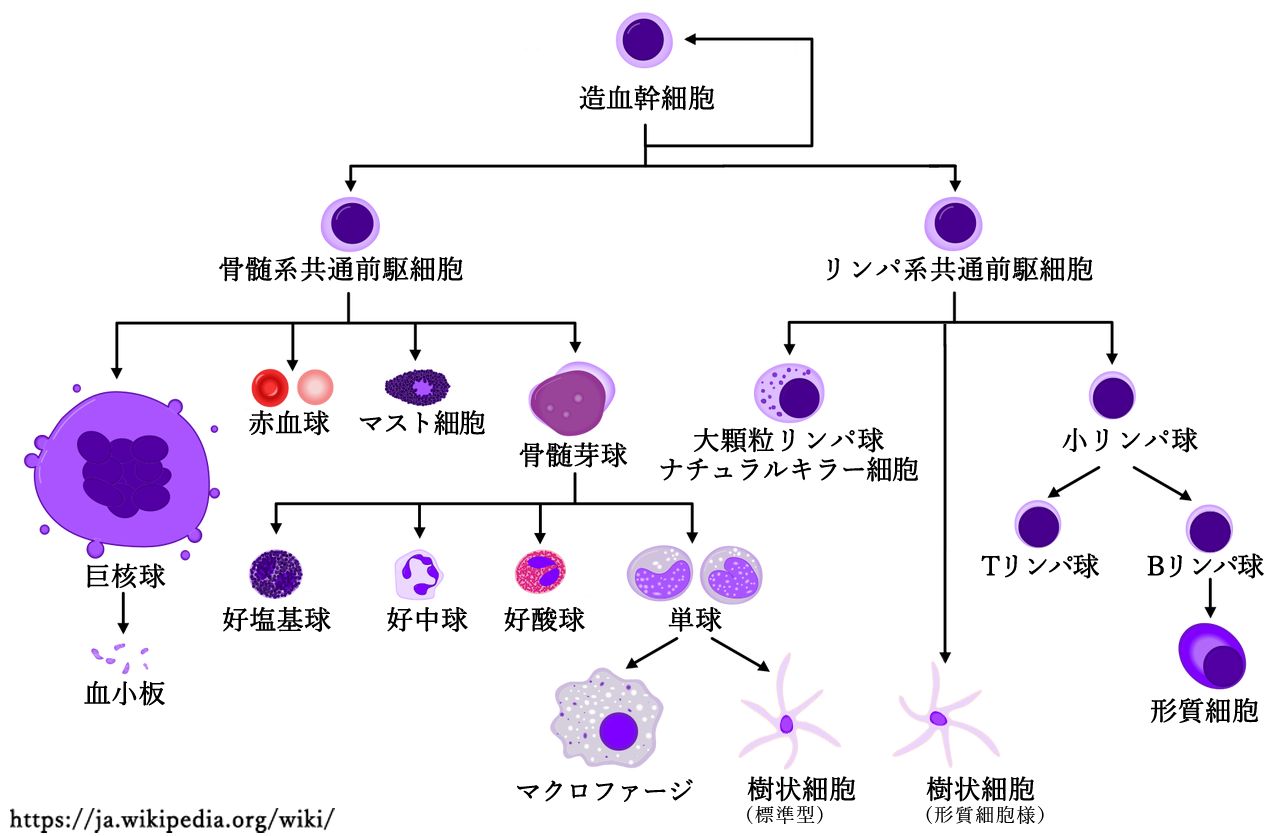
●免疫担当細胞
免疫担当細胞は 造血幹細胞由来 で,顆粒球(好中球,好酸球,好塩基球,マスト細胞)~60%,リンパ球~30%,単球-マクロファージ系細胞群~5%,樹状細胞などがある.
●抗 体;Fabドメイン と Fcドメイン
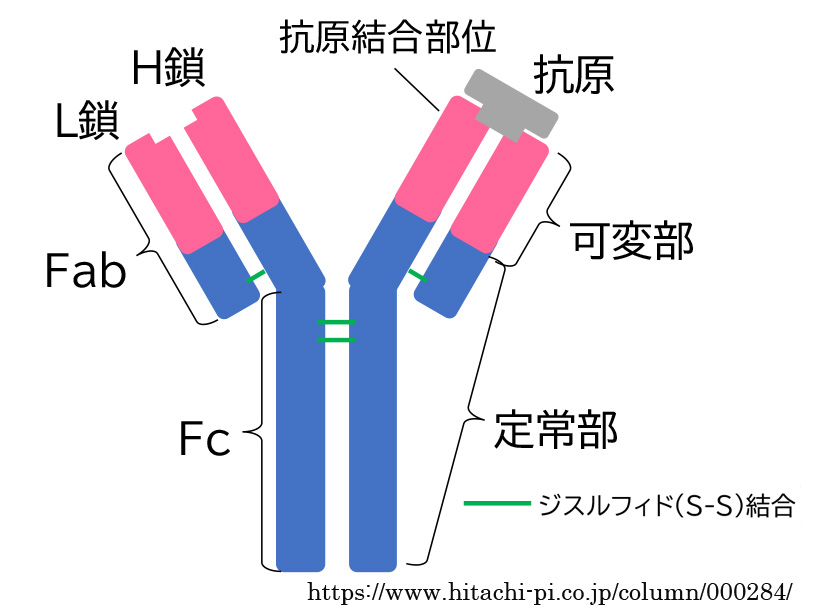
抗体は,2本のH鎖(heavy chain)とL鎖(light chain)から構成されるY字形の構造で, 「Y」の上半分の「V」の部分は抗原を認識・結合部位となるFab領域, 「Y」の下半分の「𑁇」の部分は免疫・炎症細胞などと結合するFc領域である.また,Fab領域には抗原の性質によって変化する可変部があり,定常部には抗原結合性をもたないFc領域がある. 抗体は,標的細胞の抗原に対しFab部を介して結合する.
✓▣ Fabの「F」は「断片(Fragment)」、「ab」は「抗原結合性(antigen binding)」を指し、Fcの「c」は「結晶化可能(crystallizable)」の意味である.
✓▣ 抗体依存性細胞傷害 と
Fc受容体:
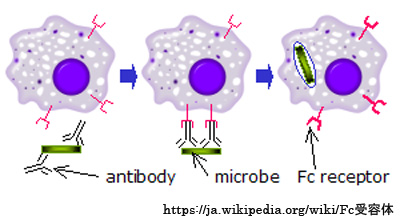
Fc受容体は,エフェクタ細胞(K細胞)の表面にあるレセプタである.抗原と結合した抗体は,免疫・炎症細胞などの細胞表面のFc受容体により定常領域で捕捉される.Fc部がエフェクタ細胞のレセプタに結合するとエフェクタ細胞内に信号が伝わり,細胞は免疫実効機能(貪食を含む細胞傷害活性や抑制)を発揮する.
Ⅱ型アレルギー(抗体依存性細胞傷害型)に関連する,細胞性免疫の一つとなる.
【 ADCC 】
●抗体の作用
✓▣ オプソニン化:微生物などの抗原に抗体や補体が結合することにより,抗原が食細胞に取り込まれやすくなる現象.
●抗体産生のストーリー
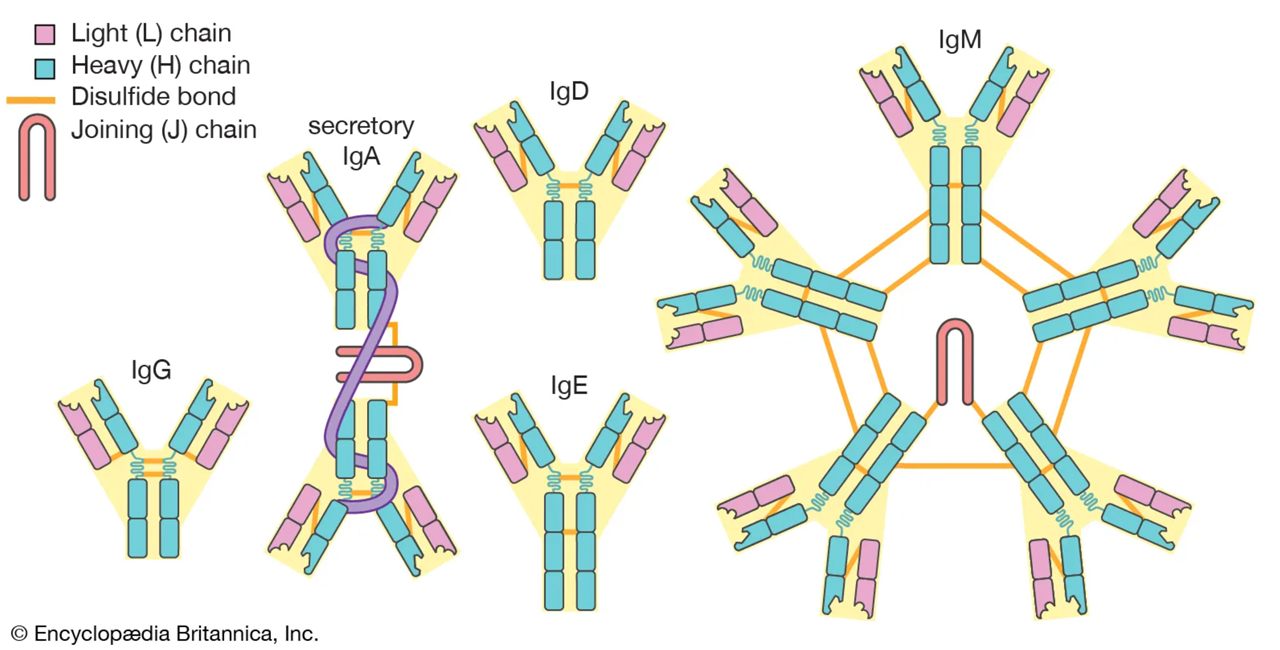
✓▣ これらにより,HLA,抗原提示,免疫グロブリンは免疫における三大要素といわれる.
●免疫グロブリン
免疫グロブリン(Ig)は抗体としての機能と構造を持つ蛋白で,抗原による免疫応答で形質細胞(plasma cell ← 元はB細胞)が産生して液性免疫に寄与している.抗体とは「感受性のある動物において、抗体の産生を引き起こした抗原と、特異的に結合することが可能な免疫グロブリン」と定義され,外来性の分子による体内への侵入に反応して産生される.
すべての抗体は免疫グロブリンであり,血漿中のγグロブリンにあたる.
クラス:IgA1,IgA2,IgD,IgE,IgG1,IgG2,IgG3,IgG4,IgM(これらをアイソタイプ isotype という)
| ・ | Ig A (Immunoglobulin A)  |
: | 局所の粘膜上皮や腺上皮の関連リンパ組織において,IgM+ B細胞からクラススイッチ組換えを介して分化するIgA産生形質細胞から産生される.血液中では主に単量体,涙液・鼻汁・気道粘液・唾液・消化管分泌液・乳汁(母子免疫)・尿中などは主に二量体(分泌型)として含まれる.侵入してきた多くの種類の病原菌やウイルスなどの侵入を防ぎ(中和抗体),粘膜面での局所免疫(粘膜免疫)に関与する.15%.体内では2番目に多い サブタイプ;IgA1,IgA2 |
| ・ | Ig D (Immunoglobulin D)  |
: | B細胞の表面に結合している.Ig M とともにB細胞上の抗原受容体として機能する.リンパ球の成熟・分裂になんらかの役割を果たしているものと考えられている.1%以下. |
| ・ | Ig E (Immunoglobulin E)  |
: | 寄生虫感染などの場合のエフェクター分子(即時反応型の皮膚感作抗体・レアギン抗体).マスト細胞や好塩基球(の高親和性IgE受容体;FcεR1)に結合し,I型アレルギー反応を惹起する.免疫グロブリンとしては最も量が少ない. Ig Eは免疫グロブリンのなかで唯一レアギン活性を持つ,即時型アレルギーに関与する抗体である.Eとは5番目に発見されたこととアレルギー皮膚反応のErythemaの双方を意味するのだとか. |
| ・ | Ig G (Immunoglobulin G)  |
: | 二次免疫応答後に産生され,細菌・ウイルス・薬物・組織抗原などの刺激に対する液性免疫の最も重要なエフェクター分子である.胎盤通過性(母子免疫)・補体活性化・貪食細胞への結合性(オプソニン化)などに優れる.血管内および血管外に存在し,5種類の免疫グロブリンのうち最も多量に存在する.75%.IgGは粘膜表面へ分泌されない。 サブタイプ;IgG1,IgG2a,IgG2b,IgG3,IgG4 |
| ・ | Ig M (Immunoglobulin M) 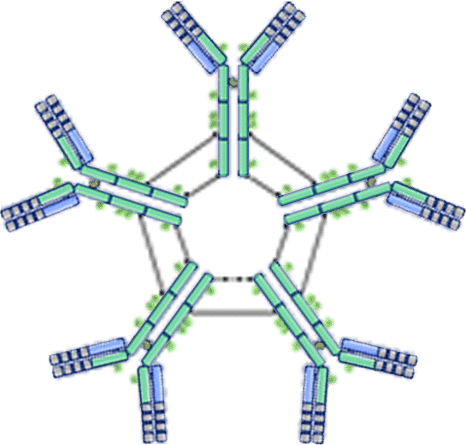 |
: | 感染に反応した時に免疫反応の初期段階(一次免疫応答)で機能する(新顔の抗原に曝露した後に最初に形成される抗体).主に血管内を循環し,B細胞の活性化・抗体産生やアポトーシス,補体活性化により病原菌やウイルスなどの抗原を破壊,白血球の食作用に関与するなどの免疫系に関与する多様な機能を持つ.10%. |
●寄生虫(蠕虫類)感染
蠕虫 helminth 感染では,血液中のIgEが上昇し,好酸球数が増加する.
虫の排出する抗原がTh2細胞やマスト細胞を活性化させ,ILⲻ4によりB細胞からIgE抗体が産生される.抗原とIgEはマスト細胞に脱顆粒を起こさせ,好酸球が虫の近くに集まる.寄生虫の周囲で,好酸球は抗体や抗原の作用で脱顆粒を起こし,リソゾーム物質・主要塩基性蛋白・活性酸素を放出して寄生虫を処理しようとする.抗体依存性細胞傷害作用(antibodyⲻdependent cellⲻmediated cytotoxicity;ADCC) の一つである.
また,感染によりATP濃度が上昇し,ATPシグナルはマスト細胞を活性化させILⲻ33を産生する.これによりICL2をスタータとする自然免疫応答が惹起される.
寄生虫感染の代表は,アニサキス症 (anisakiasis),イヌ回虫症 (toxocariasis),住血吸虫症 (schistosomiasis:日本住血吸虫) である.
●サイトカイン cytokines
細胞が産生する可溶性の低分子蛋白で,それに対するレセプターを持つ他細胞に作用し,その伝達情報により細胞の増殖・分化・機能発現などの制御を行う.すなわち生体内で,免疫・生体防御,炎症・アレルギー,発生・分化,造血機構,内分泌系・神経系などに直接あるいは間接に関与する生理活性物質で,これらによって,血管透過性亢進・平滑筋の収縮・腺分泌の亢進・好酸球の遊走などがおこる.
本来,生体防御反応であるが,時としてサイトカイン制御システムの過剰な反応あるいは変調・破綻のために,各種疾病の病態形成にも大きく関係する.
●移植免疫とHLA抗原
移植抗原はマクロファージによって T細胞に提示される.非自己タンパク抗原を認識したT細胞はILⲻ2を分泌することで細胞障害性T細胞を活性化し,抗原を持つ移植片を破壊する.これが拒絶反応である.拒絶反応とは免疫機能の一部を表している.急性拒絶反応は,T細胞が関与する細胞性免疫により3か月以内に起こる.慢性拒絶反応は,B細胞~抗体が関与する液性免疫により3か月以降に起こる.
組織適合性つまり自己抗原を決定する中心が “MHC/HLA”
●自己免疫疾患とHLA抗原
ⓐHLAⲻDR抗原の相関が指摘されている.
ⓑ
ⓒ
●疾患感受性とHLA抗原
ⓐ多様性,すなわち個人差のため疾患のかかり易さが違ってくる.
ⓑ免疫応答を直接コントロールする.
ⓒ抗原を誤認し(分子相同性),交叉反応を誘発する.
ⓓ本当の原因遺伝子座の近くにあるため,
ⓔMICA(MHC classIchainⲻrelated gene A)との関連
などが考えられる.
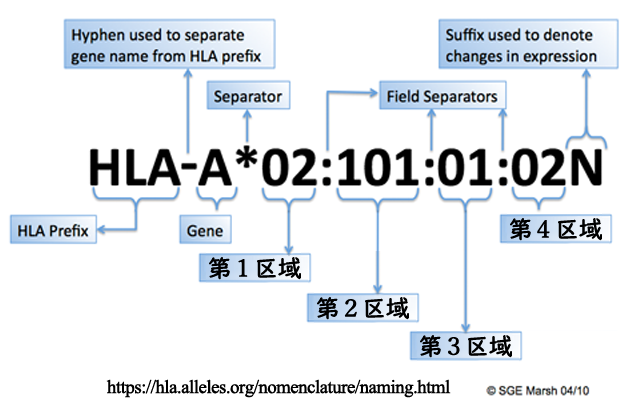
●HLAはもともとHLA抗原型として血清学的に分類されたが,遺伝子解析の進歩により,遺伝子レベルでの分類(HLA対立遺伝子型,HLAアリル)に変わってきた. 2010 年の改定(WHO 命名委員会)では,まず遺伝子名(HLAⲻA や HLAⲻDRB1 など)を記載し,その後に *(アスタリスク)を付記することで,これが遺伝子レベルでの分類であることを示すとした. 抗原型での HLAⲻA2 は遺伝子型では HLAⲻA*02 となる.さらに遺伝子レベルでの塩基配列の差異(塩基置換)による識別のため命名された区域の数字が記される.
●CD/細胞表面分子:cluster of differentiation (分化抗原群ファミリー)
CD分類は細胞表面や細胞内に発現している“抗原”の名称で,抗原(分子)に対するモノクローナル抗体を反応特異性に基づいて国際的に分類整理したものである.当初のCD番号は白血球とその前駆細胞の分化抗原を調べることを目的としており,たとえば,CD3が付いていたらT細胞,CD20が付いていたらB細胞と,特定の細胞表面抗原に結合する抗体であることを表し,機能的にも区別できる.これらは免疫系細胞に発現する細胞表面分子であり,免疫細胞-細胞間コミュニケーション,微小環境の感知,獲得免疫に重要な役割を果たす補助受容体(coⲻreceptor)分子である.
CD4抗原遺伝子は染色体12pter-12に存在しMHCクラスⅡ分子に対するレセプター,CD8抗原遺伝子は染色体2q12に存在しMHCクラスⅠ分子に対するレセプターとしての機能を持つ.
研究当初は HLDA(human leucocyte differentiation antigens;ヒト白血球分化抗原)と呼んでいたが,現在は,HCDM(human cell differentiation molecules)と改称されている.
●LMP:low molecular mass polypeptide(あるいは low molecular weight protein)
●TAP:transporterⲻassocoated with the antigen processing
✓☂ Langerhans と Langhans:
◙ Langerhans cell → ランゲルハンス細胞.単球由来で皮膚に居て,抗原提示に関連する 樹状細胞.
◙ Langhans giant cell → ラングハンス巨細胞.類上皮細胞肉芽腫に関連
2025