| 網 膜 |
| retina |
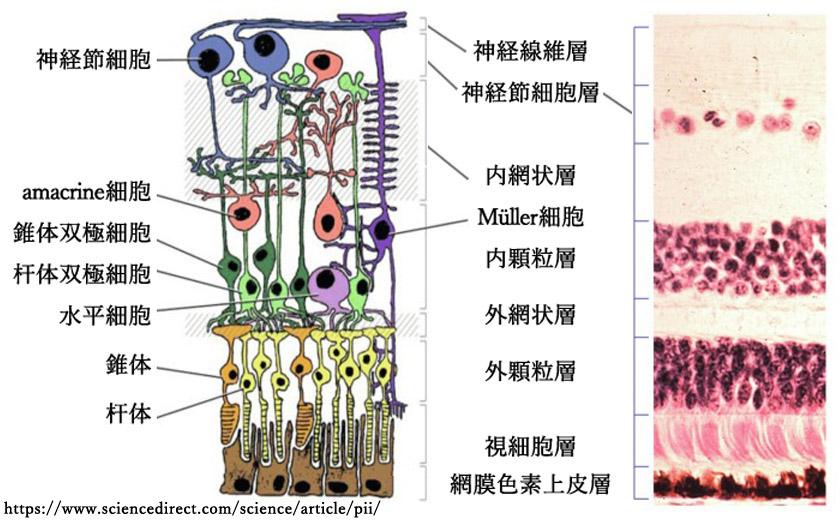
網膜は眼球壁の最内層 tunica interna bulbiにあって光を受容し,視覚処理の初めの段階を受け持つ薄い神経膜 tunica nervosaである.
我々の網膜に届いた光は,視細胞受光部分に到達するまでに毛細血管や神経細胞層(視細胞核を含めた双極細胞・神経節細胞など)を通る.視細胞層の後ろには色素細胞層があり,散乱した光を吸収して,網膜への乱反射を防止している.
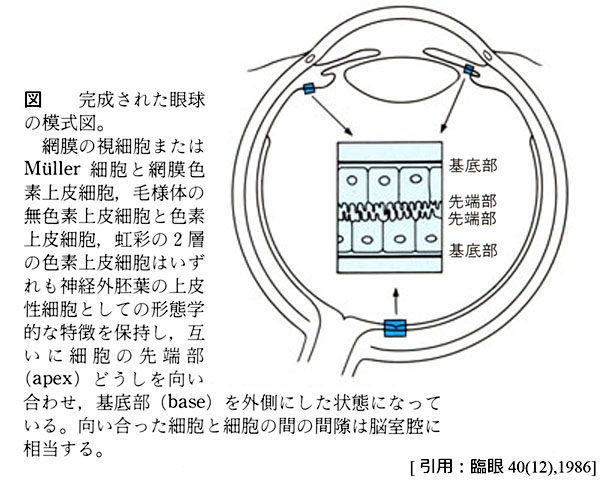
臨床でいう網膜は,視神経乳頭部 optic diskから鋸状縁 ora serrataまでの光受容部 pars optica retinaeを指している.すなわち,脈絡膜に対応する範囲である.鋸状縁で毛様体上皮に移行する.発生上,内外の二重構造になっている.
感覚網膜 sensory retina(neural retina):
眼杯内板から発生し基本的に三階層の神経細胞層で構成される.よーするに視細胞から内側.
網膜色素上皮 retinal pigment epithelium:
眼杯外板から発生し網膜としては最外層にある.
網膜は胎生早期に脳胞から伸び出た眼胞 optic vesicleを原基としている.眼胞は中央がくぼみ,二重構造の眼杯 optic cupになることで,内外の二層のあいだが “潜在的”な空間となる.網膜下腔 subretinal spaceである(よって網膜下とは 脈絡膜を意味するものではない).これにより,眼杯 → 網膜(tunica interna)= 網膜視部(pars optica)+網膜盲部(pars cæca) ということで,網膜の二重構造は毛様体・虹彩の二層の色素上皮に連続している.
網膜視部は眼球内面の65〜72%ほどをカバーする.内径22mmの球として単純計算すると,網膜面積は988〜
感覚網膜の厚みは,視神経乳頭周りで500〜600µm,黄斑周囲で250〜300µm,中心小窩で150〜200µm,鋸状縁で約100µmである.
♢ 細胞間質の少ないタイトな構造であるが,感覚網膜全体として90%以上の含水量とのことである.(https://www.ncbi.nlm.nih.gov/pmc/articles/PMC10411642/
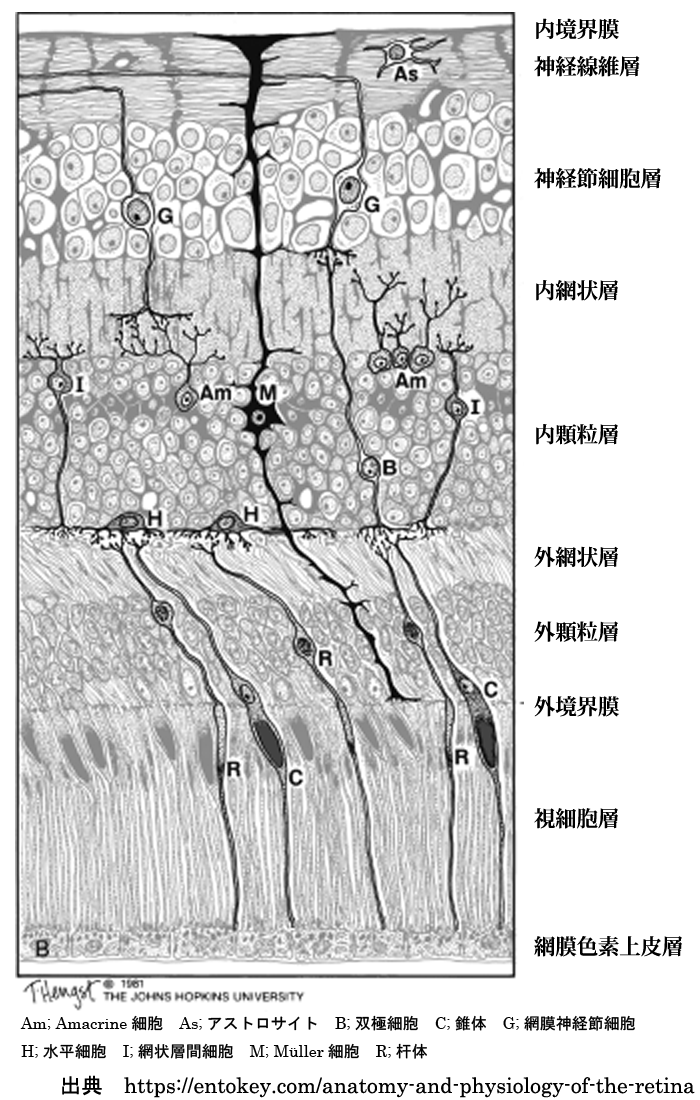
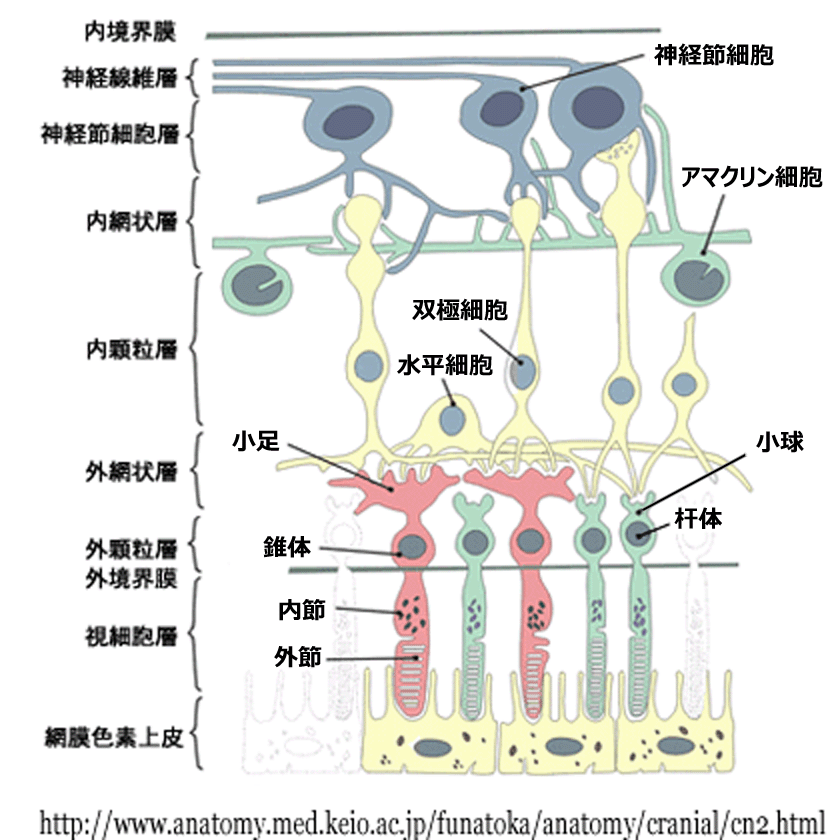
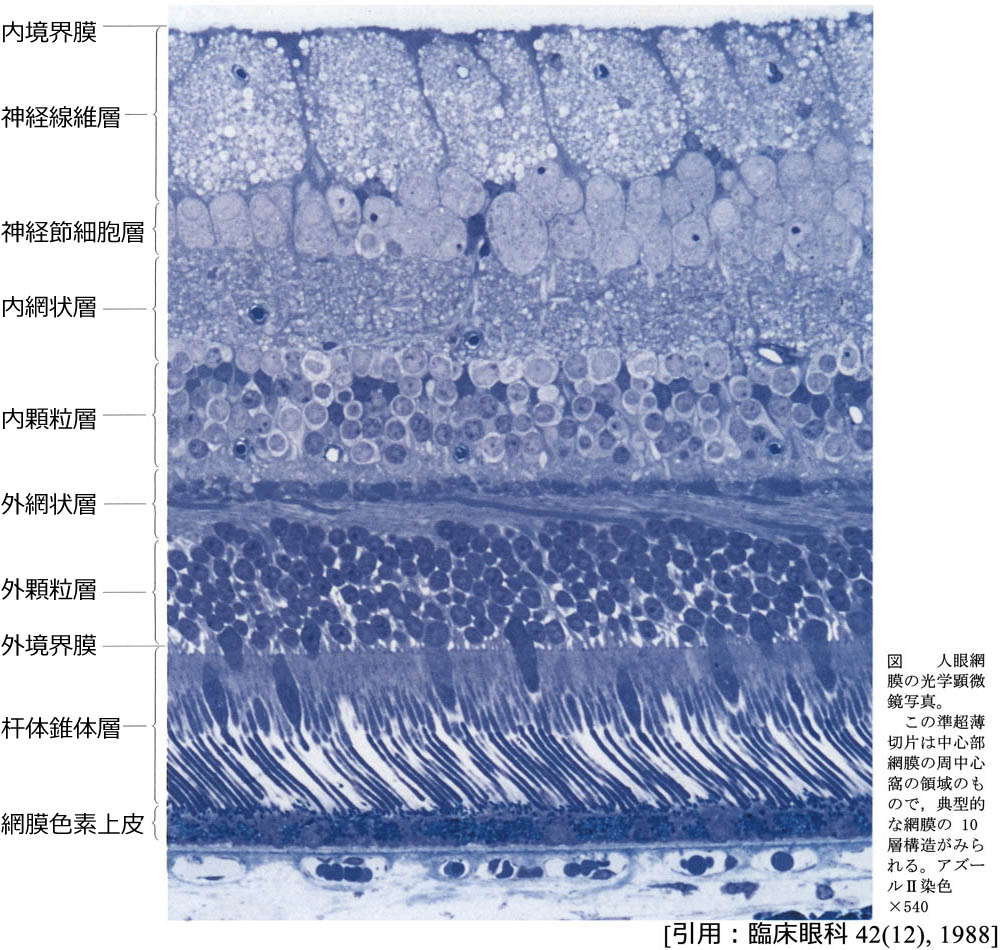
❶内境界膜 internal limiting membrane / inner limiting membrane
内境界膜
membrana limitans interna retinae/stratum limitans internumは,Müller細胞の突起(基底膜,または end feet)に,硝子体線維・ムコ多糖をまじえて形成される.
後極部・傍中心窩領域で厚く1.5⁓2.5µm,鋸状縁近くでは 約0.1µm,
硝子体に接する側はコラーゲン(IV型―基底膜成分)が付着し,硝子体コラーゲン(II型)の足場になっている.主な接着分子はフィブロネクチン・ラミニンである.
❷神経線維層 nerve fiber layer / optic nerve fiber layer
視神経線維層 stratum fibrarum nervearumは,神経節細胞の軸索の集合で,Müller細胞の突起によって小さな神経線維束に分けられている.100(⁓140)万本ほどの神経線維は視神経乳頭に集まる.アストロサイトを伴う網膜の主要血管を含む層でもある.
視神経とはこの軸索のことで,眼内では無髄軸索であるが乳頭部から篩板を抜けた眼外では有髄となり,主に外側膝状体へ達してニューロンを替える.
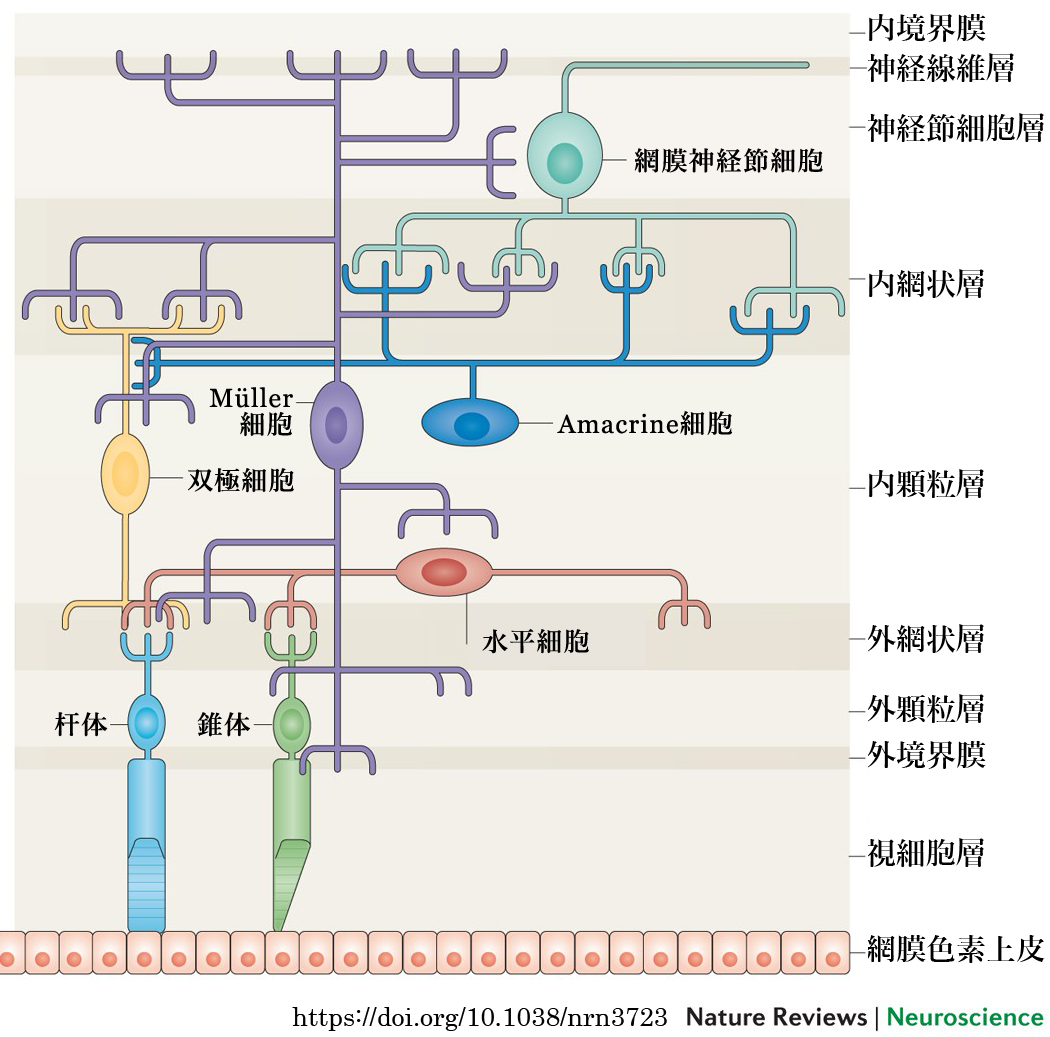
❸神経節細胞層 ganglion cell layer
神経節細胞層 stratum ganglionicum retinae/stratum ganglionare nervi opticiは,網膜神経節細胞(第二ニューロン;2nd neuron)の核(細胞体)の部分である.黄斑周囲では核が8⁓10列をなし(60⁓80µm),外側では単層となる(10⁓20µm).アストロサイトのほか異所性のamacrine細胞を含む.
❹内網状層 inner plexiform layer
内網状層 stratum plexiforme internum retinaeは,双極細胞の軸索が,神経節細胞の樹状突起や amacrine細胞の神経突起とシナプスを作る部分である.第2段階の情報処理層として並行処理を行う5⁓6層が区別できるそうである.
❺内顆粒層 inner nuclear layer
内顆粒層 stratum nucleare internum retinae/stratum granulosum internum/stratum ganglionare retinaeは,双極細胞(第一ニューロン;1st neuron)・Müller細胞・水平細胞・amacrine細胞・網状層間細胞の核(細胞体)の部分である.amacrine細胞はIPLに面している.水平細胞はOPL側にある.
❻外網状層 outer plexiform layer
外網状層
stratum plexiforme externum retinaeは,視細胞の軸索が,双極細胞の樹状突起や水平細胞の神経突起とシナプスを作る部分である.外側
⅔は視細胞体から伸びる軸索,内側
⅓はシナプスないし樹状突起.中心窩での視細胞軸索はHenle線維とよぶ特殊形状になっている.なお,
視細胞軸索は外顆粒層のパーツと見做す研究者もいる.ここまで神経上皮層に属するということだ.
❼外顆粒層 outer nuclear layer
外顆粒層 stratum nucleare externum retinae/stratum granulosum externumは,視細胞の核の部分である(核を含む細胞体perikaryaとその突起).黄斑ではわずかながら Müller細胞の核が紛れ込んでいる.中心窩では薄いHenle線維と内境界膜を介して硝子体に接している.
❽外境界膜 external limiting membrane / outer limiting membrane
外境界膜 stratum limitans externum retinae/membrana limitans externaは,Müller細胞と視細胞との結合複合体 junctional complexまたは接着帯による細胞間連結装置.Müller細胞の頂端 apex/突起部 に相当し,視細胞同士を固定する終末堤 terminal barである.穴あき網目シートで穴に視細胞をはさんでいるイメージであるが,実際の膜組織ではない.
❾視細胞層 photoreceptors layer
視細胞層 は,光感覚細胞(受容体細胞)の内節・外節 stratum segmentorum externorum et internorumの部分である.杆体rod と 錐体cone が混在して(duplex retina),一層を成している.
 |
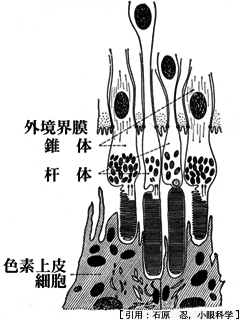 |
❿網膜色素上皮層 retinal pigment epithelial layer / stratum pigmenti retinae
色素上皮細胞RPEは一層の整然と配列した形状で六角形を示す.平常は分裂しないが種々の病的状態で分裂・増殖をきたす.基底膜の Bruch膜を介して脈絡毛細血管板につながり,グリアの機能を持つ単層立方上皮の位置づけとして機能的には脈絡膜に属する.その機能は,
血液眼関門を形成・・・・・細胞同士の tight junction による
神経網膜を接着・・・・・網膜下腔のムコ多糖類の合成,ポンプ作用
網膜外層の栄養・・・・・栄養および代謝産物の輸送
ビタミンAの代謝・・・・・ビタミンAの貯蔵と視色素の合成
視細胞外節の貪食・・・・・消化・リサイクル,リポフスチンとして蓄積.
メラニン顆粒(チロシンから作られる)による暗箱機能・・・・・散乱光吸収.メラニン色素量は黄斑領域が最も多い.神経堤由来の脈絡膜のメラノサイトと違いRPE色素は人種での違いはわずかであるとされる.
胎生期の視細胞形成に必要
免疫反応・サイトカインの分泌
VEGF分泌による脈絡膜のメンテナンス
などなど
細胞数350〜600万個(高齢者で多くなる),細胞密度は周辺で下がる(中心では5,000/mm²,周辺では2,000/mm².
眼球へ入る血管は,眼動脈 ophthalmic arteryから分かれた網膜中心動脈 central retinal arteryと毛様(体)動脈系 ciliary arteriesである.網膜への栄養は,網膜に直接分布する網膜中心動脈の枝と,毛様動脈が接続する脈絡膜による.
網膜中心動脈系の守備範囲は外網状層よりも内側,すなわち第一ニューロン以降の 脳層 である.脈絡膜の守備範囲は外網状層より外側,すなわち 神経上皮層(視細胞,網膜色素上皮細胞)の範囲である.これにより双方の境界は外網状層(視細胞と双極細胞とのシナプス)部となる(この部を
中境界膜 と言うことがある).
 |
 |
 |
 |
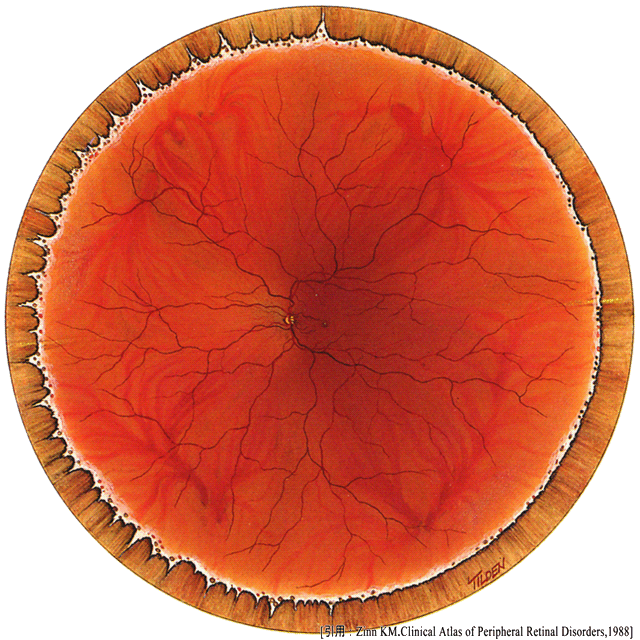
構造というと断面が要求されることが多いが,平面つまり瞳孔を通して眼球内部を見たものが眼底である.網膜とその血管,脈絡膜,視神経乳頭が眼底所見の主要な構成要素である.
眼底は平面を主体にした広さの概念であるから,部位によって機能が違い,発症疾患の差となる.
| 視神経乳頭 optic disk | … | 視細胞(というか網膜構造そのもの)を欠くため視野の中では盲点となる |
| 中心窩 fovea | … | いくつかの層が外側へ押しやられている → 光の透過性増加・結像の収差減少 毛細血管が無い,錐体外節が異常に長い,小さい受容野 → 高い解像度 |
| 後極部 posterior pole | … | 乳頭・黄斑部・血管アーケード一帯を指す |
| 赤道部 equator zone | … | 中間周辺部・渦静脈の深さ |
| 周辺部 periphery | … | 網膜は鋸状縁で終わる |
疾患臨床で『眼底所見は ?』と言うと,地図のような一定の広がり(範囲あるいは位置)や隆起のほかに,地質図のような深さ,脈絡膜の外側つまり強膜さらに眼外の病変を言外に含むことが多い.同時に硝子体側の情報も必須である.
【 ❶【第三章】脈絡膜, ❷【第五章】眼底, ❸【第十一章】血管, ❹【第十三章】発生 】
![]()
視細胞で感知された光情報は,電気情報として網膜内の神経系ネットワークで変調され,視神経では神経インパルスの形で中枢に伝えられる.ネットワークによる情報処理とは即ち脳機能であることから,網膜脳層と云われる.ということで,
網膜階層は,何通りかの分け方ができる.
㈠ 10層構造;微細構造の基本
㈡ 2層,すなわち感覚網膜+色素上皮;発生上の眼杯に由来
㈢ 2層,すなわち脳層+神経上皮;機能・血管分布に由来
㈣ 3層,すなわちニューロンの構成階層
などなど.網膜内での視覚情報の流れの基本は,
視細胞 ⇒ 双極細胞 ⇒ 神経節細胞, すなわち
„ニューロンの3階層“ で解釈される.
(光受容細胞)(第一ニューロン)(第二ニューロン)
ちなみに,視細胞:双極細胞:神経節細胞 =100:17:1 ほど,とのことである.
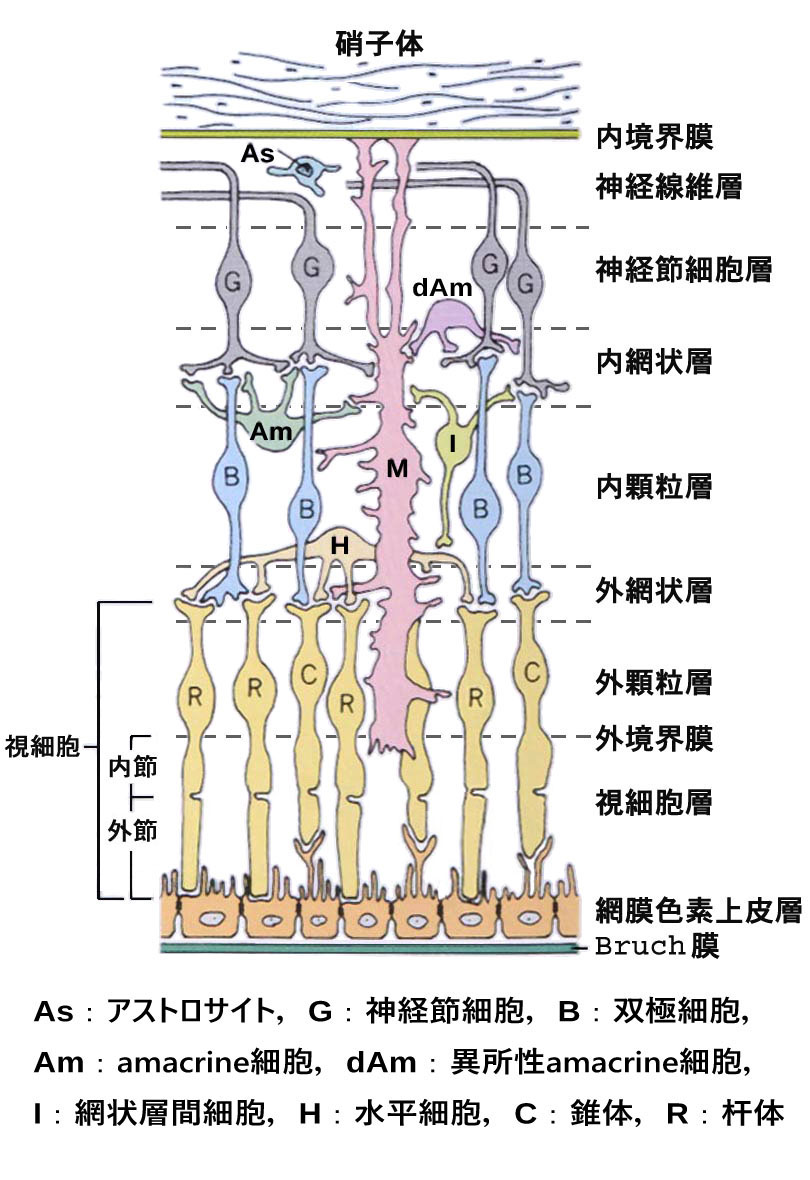
| 視細胞 photoreceptor cell | : | 杆体 rod と錐体 cone.核・細胞体は外顆粒層を形成. |
| 水平細胞 horizontal cell | : | 内顆粒層の視細胞寄りに存在. |
| 双極細胞 bipolar cell | : | 内顆粒層を形成する主要素. |
| amacrine細胞 | : | 内顆粒層の神経節細胞寄りに存在. |
| 網状層間細胞 interplexform cell | : | 内外の網状層を連結する細胞. ドーパミン作動性の特殊なamacrine. |
| 神経節細胞(視神経細胞)
retinal ganglion cell |
: | 多極神経細胞.軸索は視神経となる. |
| グリア細胞 | (glia 神経膠細胞;情報伝達には関わらない神経系細胞) |
| Müller細胞 | : | 網膜の主要グリア.核はおもに内顆粒層に存在.網膜の支柱的役割と光の誘導,神経細胞保護作用,キサントフィルの保管.網膜神経細胞と共通の前駆細胞から作られる. |
| 網膜色素上皮細胞 | : | Müller細胞 と共に,細胞内外のKイオン濃度の変化により膜電位が変化する生理特性がある. 膠細胞と同等の機能がある,と説明される. |
そのたのグリア 【 ✍ グリア 】
| ✩星状膠細胞(astroglia,astrocyte):網膜血管周囲・乳頭表面に存在するグリア.視神経を介して網膜内へ移動してくる.栄養・関門機能. |
| ✩希突起膠細胞(oligodendroglia,oligodendrocyte):視神経で髄鞘を形成. |
| ✩小膠細胞(microglia):内網状層に存在.貪食と組織修復,神経保護.中胚葉性 primitive macrophage 起源. |
| 血管系の細胞 | : | 内皮細胞,周皮細胞(壁細胞),平滑筋細胞,線維芽細胞,など. |
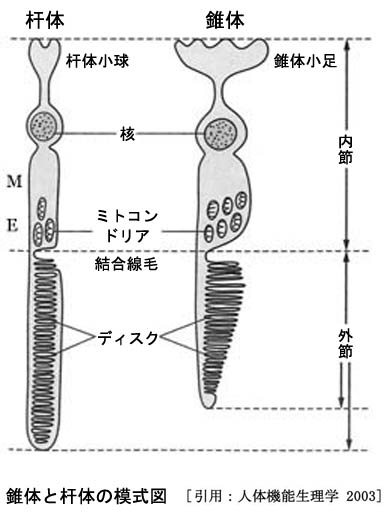
視細胞は,神経外胚葉から発生した神経上皮細胞が光感覚細胞として機能するようになったものである.光受容部が外節 outer segmentで,視色素はディスク(円板膜)に付着する.結合線毛 connecting ciliumを介し内節 inner segmentと接続している.内節は細胞内器官に富み,外節側はミトコンドリアを多く含む部分でエリプソイド(ellipsoid;楕円体部
線毛(繊毛)が分化した外節は,刻々代謝され日々10%ほど入れ変わっている.細胞分裂をしない視細胞として,光フリーラジカルによるダメージを解決するしかけになっている.線毛は,感覚細胞として外界のシグナルを受け取るアンテナのような役割を果たしていたものである.線毛細胞の一種として原始生物時代の名残りの構造をとどめているのだとか(線毛型光受容細胞).これにより,(非運動性)線毛は結合部 connecting pieceと呼ぶのが一般的である.
軸索の長さは錐体・杆体では差がなく,場所により差があるとのことで最長5〜600µm.
シナプス部では双極細胞の突起が1本と水平細胞の突起が2本の計3本が一組となって視細胞の軸索を受ける.シナプス結節 synaptic terminalは,錐体・杆体でそれぞれ呼び名が変わる(図).
なお引用図での‛内節’は軸索部を含めた説明となっている.内節はテキストにより,軸索部以外を,あるいはミオイド部とエリプソイド部を指したりする.機能的には‛M+E’が妥当かもしれない.外境界膜部の外側ということになる.
⏩ 基は神経細胞:すなわち外網状層のシナプス結節の軸索突起(視細胞内線維)に対し,網膜下腔(眼杯腔)に向かう内節・外節は樹状突起に相当する.視細胞外線維は,細胞体から外境界膜までの部分で,樹状突起の一部となる.
⏩ ミオイド:明⇄暗で伸び⇄縮みする ?
⏩ 結合線毛:外節は結合線毛から発生する.視色素など外節の部品は内節で合成され,線毛部を通して外節円板に輸送される.線毛部を外節基底部,円板部を外節辺縁部ということがある.
線毛の機能障害は,網膜色素変性を合併する先天全身疾患に関連する.
【 ✍
繊 毛 】
⏩ 繊毛型光受容細胞:光により過分極応答.脊椎動物の繊毛由来受光部(視細胞)の反応.
感桿型光受容細胞:光により脱分極応答.無脊椎動物の微絨毛由来受光部の反応.メラノプシン神経節細胞も同じ.
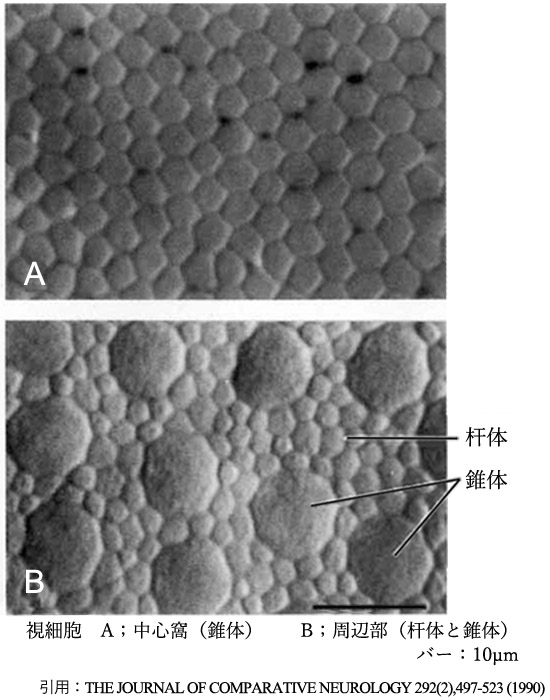
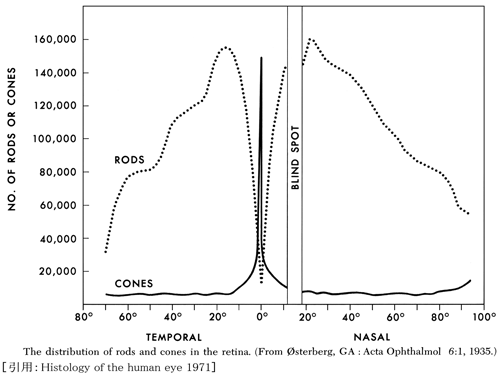
❶ 錐体 cone:約500万個(350⁓700万個).内節径は2〜6µm(中心窩➠周辺)で長さは75µm.基本的に黄斑に分布し,数個の錐体が1個の双極細胞に収束するため高解像度で,特に中心窩では神経節細胞まで1対1対1対応による最高分解能を得る.感度が低く(閾値が高く)明所で機能する. これにより明所視での図形の形状識別(または認識≒視力)をしている.径は特に中心小窩でスリムになっているが,高密度になるには成人までかかる.
外節(形質膜の折畳みからなる円板膜;錐体では露出)が短く,ラメラ膜という.外節の脱落は明所視→暗所視に変化した直後に増加するとのことである.
錐体細胞数は中心小窩の領域で200,000個/mm²(10〜30万個/mm² と,中心小窩の範囲というか研究者の報告によって幅がある.また,錐体サイズを2〜2.5µmとすると単純計算で25〜16万個/mm² となる).
中心小窩は凡そ ⌀1°.径350µmとすると面積約0.1mm²(径400µmで面積約0.13mm²)で,16,000〜
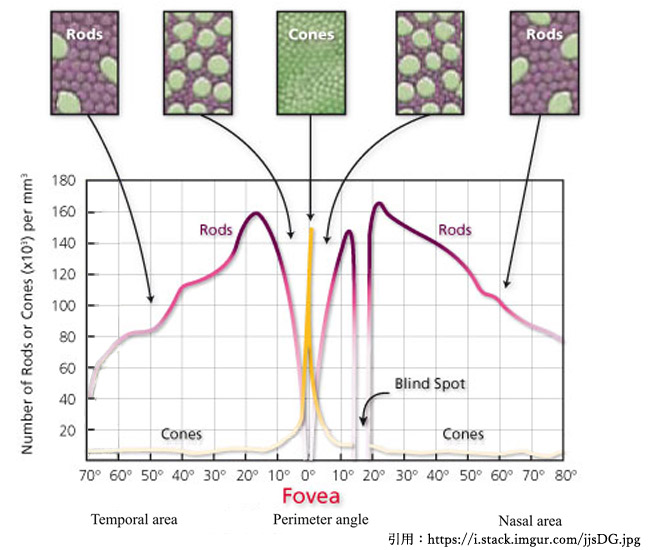
錐体数は中心から500µm離れると ½ ,10°(3mm)離れると ¼,4mm離れると 1/10以下になる(当然,杆体と混在).
中心窩は凡そ ⌀5°,径1,500〜1,750µm(1.77〜2.4mm²),100,000個/mm² ということで中心窩内には約20万個の錐体が存在することになる.
なお,凡そ 径6mm╱20°の黄斑部でも,錐体細胞数は全体の10%ほどの分布量とのことである.
⏩ 中心窩での錐体サイズを2µmとして, 2点弁別に必要な刺激されない1個の錐体を挟むと2点間は4µm, 視角は0.8′(計算の根拠)に相当する.つまり,単純計算では視力1.25が限界となる. 【 追加事項 】
❷ 杆体 rod:約1億個(7500万⁓1.25億個).径1〜2µmほど,長さは120µmほどである.基本的に周辺に分布し,感度が高く(閾値が低く)暗所で機能する.多数の杆体が1個の双極細胞に収束し(次元圧縮,というらしい)感度を上げているが,解像度は期待できない.これらにより光覚(あかるさ・光の存在の検出)・動覚にかかわる.
外節(rhodopsin分子を豊富に含む円板が自由に浮遊)が長く,ディスク膜という.外節の脱落は暗所視→明所視に変化した直後に増加するとのことである.
中心小窩には存在しない(rod free zone).報告者により250〜750µmと差があるが,無杆体部分は径1〜1.25°≒径350〜375µmくらいの範囲になるようである.最も高密度(160,000個/mm²)となるのは20°〜25°
杆体視色素は光量子1個に反応し,錐体の1,000倍以上の感度がある(閾値が低い).明所では動作を停止する(飽和する,という).
⏩ rod free zoneにより,暗所視では中心暗点として自覚される.
⏩ 眼軸が伸びると(23mm → 27mm)後極部の視細胞密度は80%になる.
❸錐体より杆体の方が脆弱である.疾患の影響はもとより,加齢による減少も先行する.
錐体のグルコース代謝は,杆体に依存する(RdCVF;rod derived cone viability factor 杆体由来錐体生存因子)とのことである.
❹錐体系は,瞳孔中心を通って入射する光に最もよく応答する指向性がある.→ Stiles᠆Crawford効果
杆体系は,入射部位とは無関係に,入射する光量により応答の強さが左右される.
⏩ 視細胞が隙間なく配列されているとして,形態は多角形になるらしい.模式的に六角形として計算できることになるが,
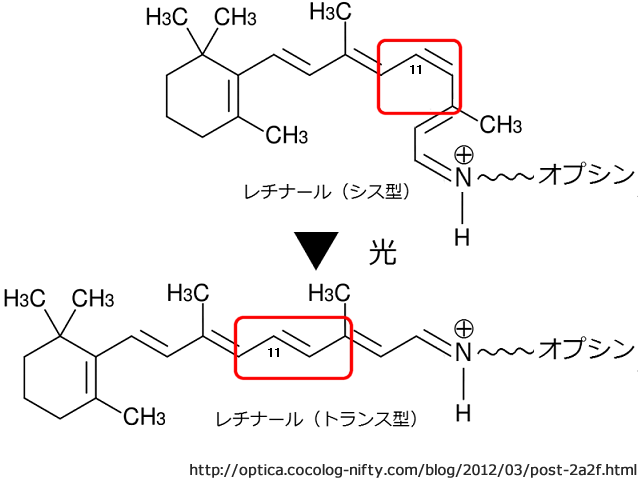
外節にあるディスク上の視物質(視色素)で光量子を受容(光を検出)する.
視物質は,vitamin A 誘導体(レチノール)のアルデヒドであるレチナール retinalを発色基(発色団)として,オプシン opsinタンパクとの複合体 retinal-opsin complexになっている.11-cis型retinalは光が当たる(色素が光を吸収する)と,全trans型への異性化で脱色(褪色 bleaching)し,opsin
と解離する.そのごの一連の反応で膜電位が変動する(受容器電位を生じる).
分離した 全trans型retinalは網膜色素上皮細胞内で(錐体のretinalはMüller細胞内でも) 11-cis型に戻り,視色素としてリサイクルされる.
光感覚のトリガーは,レチナールの異性化(電子遷移・エネルギー変換)と視色素の分解ということである.
レチナールは視色素に共通で単体での吸収域は紫外にあり,オプシンによって吸収波長を変えているとのこと.基本的に視細胞オプシンはアミノ酸配列の違う四種類があることになる.錐体オプシンはフォトプシン photopsinⅠ〜Ⅲ,杆体オプシンはスコトプシン scotopsinである.
●退色型光受容タンパク
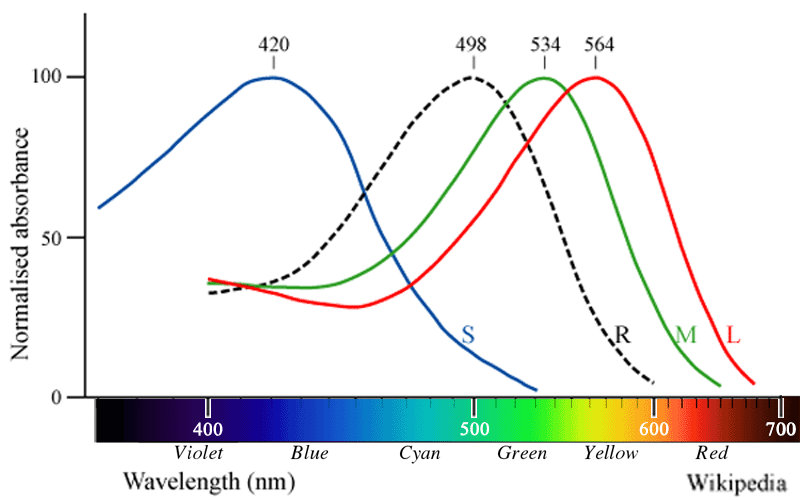
❶ 錐体は,波長感受(分光感度)特性の違う三種類の視物質(iodopsins ヨドプシン:青色素
| 短波長感受性錐体(S-cone,青錐体) | : | 最大吸収波長 419~445 nm(blue ;short wave) |
| 中波長感受性錐体(M-cone,緑錐体) | : | 最大吸収波長 530~545 nm(green ;middle wave) |
| 長波長感受性錐体(L-cone,赤錐体) | : | 最大吸収波長 557~570 nm(red ;long wave) |
分布はディスプレイ画面のような規則性・平均性はなく,結構ランダムにかたまり(群)を作ってちらばってる,とのことで,色のコントラストによる空間分解能は低い(この生理特性を利用したのが NTSCカラー伝送 ?).
錐体数は L>M>Sで,おおよそ L錐体は64%ほど,M錐体は31%ほどの割合とのことである.L:Mについては個人差が大きく,1:1〜17:1 と遺伝子上の変異を生じやすく比率はバラつくのが実際らしい.
S錐体は2〜数%ほど.中心小窩内の径100µm(0.35°)部分は S錐体が無い(M・L錐体のみの部分が 中心小窩 foveola,との解釈もある). 以前より,非常に小さい対象に対して青色盲を示すことが心理物理的に確認されていて,青色に対する最高感度は中心から1°離れた部位である.これによりS錐体による形態覚は貧弱である.ただし S錐体は周辺までほどほどの割合で分布している(とのことで青指標の色視野が広いことの説明になる,かもしれない ?).⏩ 二種類の L錐体:アミノ酸の違いにより最大吸収波長 556.7 nmのヒトが62%,552.4 nmのヒトが38%.
⏩ 個々の視色素は波長ごとに反応強度(吸収比率)が違い,比率に従って 色覚 を生じる.要するに色覚とは二種以上の視色素の相対的強度により波長を弁別することである.念を押すが,波長に “色” は付いていない.
⏩ 波長特性は,実験方法により微妙に変わる.生体眼では角膜表面上での波長特性を示し,水晶体・硝子体や黄斑色素の影響を受ける.視細胞表面での波長特性の報告では,紫外領域にも感受性があることで短波長方向へシフトした特性となっている.
👓
„blue cone“ と „blue amplifier“:
S錐体は低酸素環境に脆弱であるらしい.S錐体ERGによる研究では,網膜色素変性や糖尿病網膜症において,早期からS錐体ERG振幅が有意に減少する,また網膜剝離では復位後に於てもS錐体ERG振幅は回復しない,とのことである.ただ記録法についての完成度も研究途上なんだとか.
❷ 杆体は,一種類の視物質(rhodopsin ロドプシン)にて光吸収を行なう.最大吸収波長は500(496〜498)nm付近である.これは地球上で観察される太陽光線の波長分布のピーク(500~550 nm)と一致する.太陽表面温度の約5,800Kに依る,とのことで,表面温度の違う恒星に属する惑星に視覚をもつ生物がいるとすると,可視光線として利用する電磁波の波長は違うと考えられる.なお,自然光の波長成分については,メラノプシン神経節細胞を研究する立場でも重要視されている.
rhodopsin は,量が多く信号増幅機能があり光閾値が低いが,明順応下では信号が飽和し,明視野では錐体の抑制により機能を停止する.明順応状態にならない,あるいは錐体が抑制しない状態が昼盲となる.実際には,錐体視の状態でも杆体中の rhodopsin は 1 %しか退色しない.rhodopsin の濃度が10%下がると視感度は1⁄100になるという.
また,暗順応下の杆体は錐体を 抑制 する.
⏩ カエルの杆体は2種類:元々の緑感受性杆体視物質と青感受性錐体視物質を高感度(ローノイズ)に進化させた2種類,とのこと.すなわち,暗所視でも色覚が生じている!
❸ 視細胞(3種類の錐体と杆体)は光の波長によって感度が異なる.短波長感受性錐体(S錐体)は419~420nm付近,中波長感受性錐体(M錐体)は
対応する色はおおよそ,
420nm→紫,500nm→緑青,530nm→緑,560nm→黄緑である.
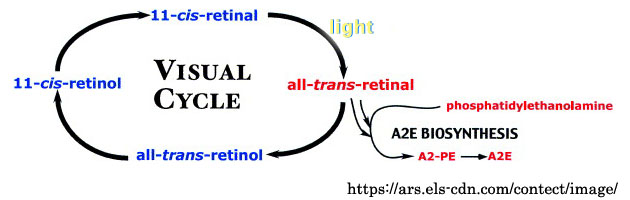
❹ 視細胞外節に存在する視物質は光を吸収するとオプシンと全trans᠆レチナールに分解される.網膜色素上皮細胞は全trans᠆レチナールが含まれる脱落した視細胞外節を貪食し,視物質の構成成分である11᠆cis᠆レチナールを再生する.一個の網膜色素上皮細胞は一日あたり約5,000枚の外節円板を貪食する,とのことである.
この視物質の代謝は視サイクル(visual cycle)またはレチノイド代謝(retinoid cycle)と呼ばれる.この過程で全trans᠆レチナールとリン脂質が反応することによりA2Eが生合成される. 【 追加事項 】
A2Eは細胞毒性があり,リソソーム膜の破壊,フリーラジカルの産生,酸化反応の亢進などにより,最終的に網膜色素上皮細胞へのダメージをもたらし細胞は消滅する.また,炎症を惹起して血管新生を促進する.また,リポフスチン の主要な構成成分で自発蛍光を発し,網膜色素上皮細胞の加齢変化やStargart病に代表される網膜変性の原因として注目されている.
●A2Eの同義語
pyridinium bis-retinoid(ピリジニウム ビスレチノイド)
di-retinoid-pyridinium ethanolamine(ジ᠆レチノイド᠆ピリジニウムエタノールアミン)
N-retinylidene-N-retinylethanolamine(N᠆レチニリデン᠆N᠆レチニルエタノールアミン)
●オプシン opsinは G蛋白共役受容体(GPCR:G protein coupled receptor)の一種 【 ☟ 追加事項 】
光を受容すると構造変化を起こし,GTP結合タンパク(Gタンパク)を介して細胞内シグナル伝達系を駆動する.
●レチナール retinalは体内で脂質であるレチノール(vit A)から産生される.
11-cisレチナールは視色素が光を受容するために必須の分子である.11-cisレチナールがオプシンと結合した光受容待機状態では,Gタンパク質活性化能が強く抑制される.一方,光を受容して全trans型に異性化すると,Gタンパク質を活性化する状態になる.
❺ L錐体・M錐体視物質の遺伝子はX染色体上(q28)にある.S錐体視物質の遺伝子は第7染色体(q22‐qter),杆体視物質の遺伝子は第3染色体(q21‐q24)にある.
●ロドプシン rhodopsinはバラ色だそうです.ギリシャ語 rhódon (ῥόδον)から
●ヨドプシン iodopsinはスミレ色だそうです.ギリシャ語 iode (īṓdēs),元のサビ色から派生・転用
プルキンエ効果:Purkinje effect
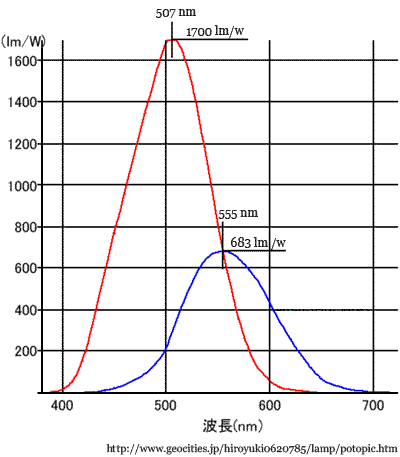
明所視では黄緑色(555nm)に最大感度があるが,暗所視では青緑色(507nm)で最も感度がよい.錐体視では黄色付近が明るく感じ,杆体視では緑色付近が最も明るく感じる,というように説明される.
目が順応できる程度にゆっくりと照明を暗くしていった時,赤やオレンジ色などの長波長に対する感度は紫や青の短波長の感度よりも早く減衰していくことになる.この
視感度 の移動shift(明⇆暗)を Purkinje現象という.
それぞれを 560nm,510nmとする教科書もある.
![]() Purkinje はドイツ語表記で,元はチェコ語で Purkyně と綴り,「プルキニェ」とするのが原語発音に最も近いのだそう.
Purkinje はドイツ語表記で,元はチェコ語で Purkyně と綴り,「プルキニェ」とするのが原語発音に最も近いのだそう.
初期の日眼用語もそんな風に書かれていたような なかったような.フルネームは Jan Evangelista Purkyně.
順応とは視物質の分解・再生のプロセスによる利得調節である.
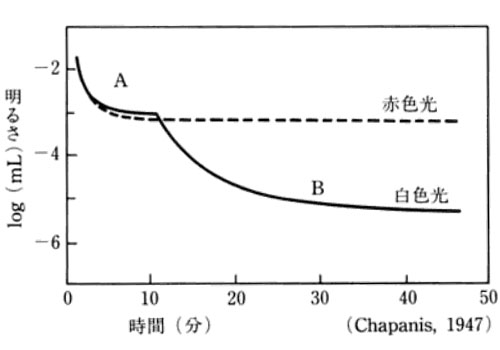
視環境の明るさは108倍で変化する.順応とは,視環境の明るさに追従して感度(閾値)が動的に変化することである.
暗い環境で光覚閾値が次第に低下することが 暗順応 となる.明るい場所から暗闇に急に入ると,始めは何もみえないが時間がたつと次第に見えてくる.この30分から1時間の過程は次のように考えられる.
光照射下では rhodopsinの再生と分解は平衡状態あるいは消費状態にある.この状態で暗闇になると分解過程が抑えられる.再生過程はそのままであるから視細胞内の rhodopsinの量が増加する.そのため,感度が良くなる(閾値が低下する).
暗順応時の時間経過による光覚閾値の低下を示した曲線が暗順応曲線 dark adaptation curveである.暗順応曲線は網膜の部位で異なる.中心窩では,明から暗に移って 2~3分以内に急速に閾値が低下し,5~10分で一定値に達する.網膜周辺部では,閾値の低下は比較的ゆっくりと生じる(30~60分).従って,暗順応曲線は二相に分かれた曲線となる.第1相は錐体,第2相は杆体が関与した過程とみられる.
暗順応 … 視物質の再生 → 視感度の上昇
明順応 … 視物質の分解 → 視感度の低下
明るい環境に変わった時,光覚閾値が上がりまぶしさを感じ難くなる状態が 明順応 となる.光への反応性が低下するこの過程は,数分で急速に完了する.錐体視物質よりも杆体視物質が容易に褪色 bleachする ?
![]() 赤色光による暗順応のデータでは,一次曲線のみ,すなわち杆体は赤色光に感度がないことを示している.
赤色光による暗順応のデータでは,一次曲線のみ,すなわち杆体は赤色光に感度がないことを示している.
眼という光感受器官の中で,受容器が視細胞,視神経で信号伝送するための変調回路が網膜脳層,ということで光を情報源とする感覚が視覚であった.視覚とは光覚・運動覚・形態覚・色覚等を包括する感覚である.
視細胞は transducer 〜変換器(光電変換)であり,誤解を恐れずに竅って言えば,視細胞もしくは視色素4種類の違いは,波長ごとの感度差(による信号強度の比)だけである.
![]() 脳層での processing 〜情報加工処理を経て漸く,錐体は明所視・形態覚・色覚にかかわり,杆体は暗所視・運動覚・光覚にかかわることになる.感覚の認識は脳に依るのは言うまでもない.
脳層での processing 〜情報加工処理を経て漸く,錐体は明所視・形態覚・色覚にかかわり,杆体は暗所視・運動覚・光覚にかかわることになる.感覚の認識は脳に依るのは言うまでもない.
網膜での主要な視覚情報の流れは視細胞 → 双極細胞 → 神経節細胞である.
光刺激は視細胞に “受容器電位”を発生させる.この電気信号は明るさの強度を情報としている.双極細胞への伝達情報の際は,水平細胞が色や明暗のコントラスト調整に関わる.双極細胞から神経節細胞への伝達情報では,amacrine細胞が再度コントラスト調整を掛ける(介在ニューロン).神経節細胞では活動電位が生じ,その軸索にそって上位の中枢(→外側膝状体→大脳)に情報が送られる.この軸索の束が視神経である.神経節細胞の軸索は主に外側膝状体へ届き,シナプスを形成している.
<網膜回路網> → → → → → → → → → 神経節細胞(G) → 大脳(主経路) ↑ 視細胞(R) → 双極細胞(B) → amacrine 細胞(A) → 神経節細胞(G) → 大脳(間接的経路) ↑ ↑↓ 視細胞(R) → 水平細胞(H) → amacrine 細胞(A) → 神経節細胞(G) ↑ ↑↓ → 大脳 (網状層間細胞(I))・amacrine 細胞(A) → 神経節細胞(G)
視細胞は光電変換をするセンサーである.視細胞内では視物質に光が当たると一連の生体化学反応 phototransduction cascadeが起こり,光覚情報を増幅し電気情報に変換する.
暗所での視細胞は 脱分極状態 にある.網膜に光が来ない状態ではグルタミン酸が常に放出されており,抑制性シナプス後電位(IPSP:inhibitory postsynaptic potential)を発生し,視神経へのシグナルは抑制されている.
明所で光エネルギー粒子(フォトン photon)が当たると視物質が分解され,cGMP-gated Na+channel はストップし Na+の流入が阻害され,細胞内の電位はますますマイナスになる,すなわち 過分極 となる.これが視細胞の電位変動である.過分極によってグルタミン酸の放出は止まり,興奮性シナプス後電位( EPSP:excitatory postsynaptic potential)が発生しシグナルが伝達される.受容した光の量が多いほど,大きな過分極がおこる.
これらにより,視物質(視色素)の光量子吸収による opsin と retinal との解離を,光覚の出発点と見做すことができる.電気情報の本体は膜電位の 過分極応答 である.視細胞電位を生じる,という表現も用いられる.
| 視細胞 ← 光情報 |
 |
| ↓ | |
| 光の照射によって過分極応答(電気情報へ変換) | |
| ↓ | |
| 伝達物質(グルタミン酸)放出の抑制 | |
| ↓ | |
| 内顆粒層の双極細胞と水平細胞へ電気情報が伝達 |
暗所では細胞内の Ca2+濃度は高い.Ca2+はS-モジュリン(リカバリン)に作用して視物質のリン酸化酵素(ロドプシンキナーゼ(RK)活性を阻止する.これにより視物質の分解が進行せず(結果,cGMPの 5'-GMPへの転換が進まず),過分極にならない(高感度状態,または入力待ち).
光の入力(過分極)によって Caチャンネルが閉じるので,明るいほど細胞内の Ca2+濃度は低くなる.それだけcGMPが消費されるので,光への反応性が低下する(感度調節,順応).錐体は反応が早いことで増幅度が低く低感度となっているのと,回復も早いことで順応が短時間となっている.明所視での杆体は入力オーバー即ち „飽和“ することで,反応性が停止する.
次の光入力に備えるには,
⏩ 光暴露により視細胞内の Ca2+濃度が低下する.グアニル酸シクラーゼが活性化する.これにより,cGMPが増産される.同時にPDEを抑制する.cGMPのNaチャンネル親和性が高まる. → 陽イオンチャンネルが開く.
⏩ 活性化&光退色した視物質のリン酸化&アレスチンの結合で不活化(過分極の停止) → 分解・脱リン酸化 → (網膜色素上皮内では)レチノールの還元 → 外節へリサイクル・視物質の再合成,の過程をとる.
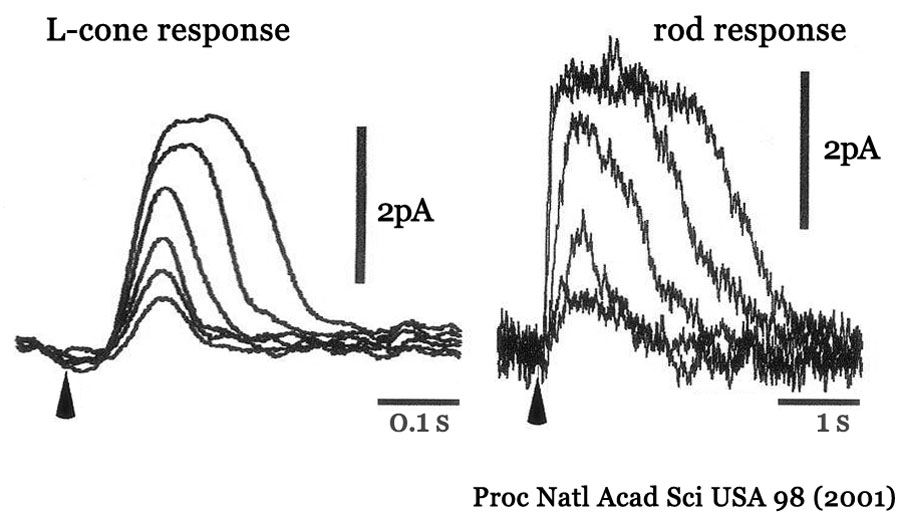
✰ 光吸収 → 過分極まで,化学反応を経由するわずかなタイムラグがあるのだそう.それでも,
錐体は応答が早く持続時間が短いことで,時間分解能は高い.杆体は応答が遅く持続が長いことで時間分解能は低い.そんなことで,視覚情報伝送は標本化された不連続信号,ということだ.
ただ,この先は受容野やら複数チャンネルやら,
コト
は簡単ではない.
✰ グルタミン酸:興奮性伝達物質
✩ リボンシナプス:伝達物質放出のため表面積をかせぐ 特殊構造 とされている.
✰ ロドプシンキナーゼ(RK):Ca依存性
✰ L錐体・M錐体のアレスチンとS錐体・杆体のアレスチンでは抗原性が違う,とのことである.
✰ 活動電位:Na+スパイク
✰ 網膜の代表的な入出力関係は,光刺激の点灯 ON と消失 OFF に対する応答である.
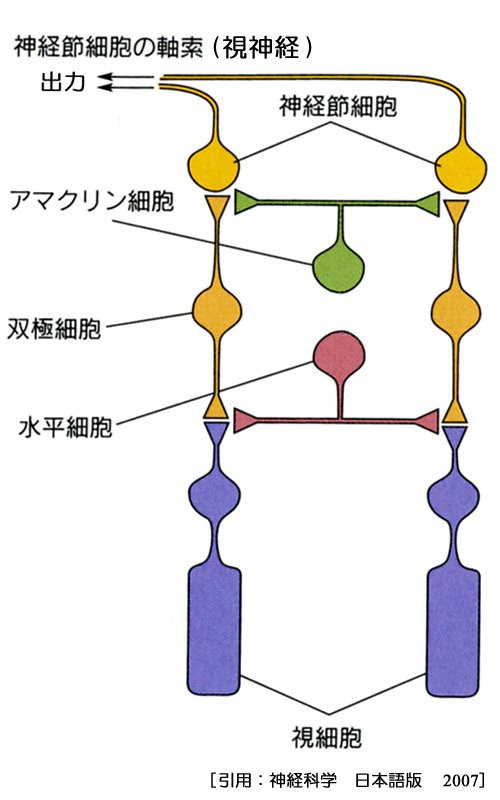
視細胞と神経節細胞をつなぐのが介在(interconnecting)ニューロンである.介在ニューロンは,双極細胞,水平細胞,amacrine細胞,網状層間細胞である.双極細胞は縦方向に連絡し,水平細胞・amacrine細胞は横方向の連絡である.
☆水平細胞:
水平細胞は視細胞とシナプスを形成し,GABA
(γⲻアミノ酪酸)
を伝達物質とする抑制性ニューロンである.
GABAは双極細胞や視細胞を過分極して周囲の双極細胞・視細胞に側抑制をかける.すなわち,暗時には視細胞からのグルタミン酸で,脱分極性の応答を起こす.光照射によりグルタミン酸が減ると過分極になる.水平細胞が過分極するとGABAの放出は抑制され,シナプス後ニューロンは脱分極する.このことによって側方抑制をして周辺受容野を形成する.
水平細胞は複数の視細胞からの入力を受けることと水平細胞同士のジャンクションにより,受容野は大型の円形となっている.暗順応時には入力の加算をして背景ノイズを減らしているらしい.明順応時には横の連絡を切り離して分解能・コントラストを確保する.
・形態的にH1・H2・H3が区別でき,それぞれシナプスしているLMS錐体・杆体の組み合わせが異なっている.1本の長い突起と数本の短い突起があり,いずれも軸索突起と樹状突起の機能を兼用する.
・外網状層では視覚の空間的情報処理を行う.
☆双極細胞:
視細胞・水平細胞は双極細胞とシナプス結合をしている.双極細胞の受容野中心部は視細胞からの直接入力,周辺部は水平細胞からの抑制入力により生成されると考えられている.
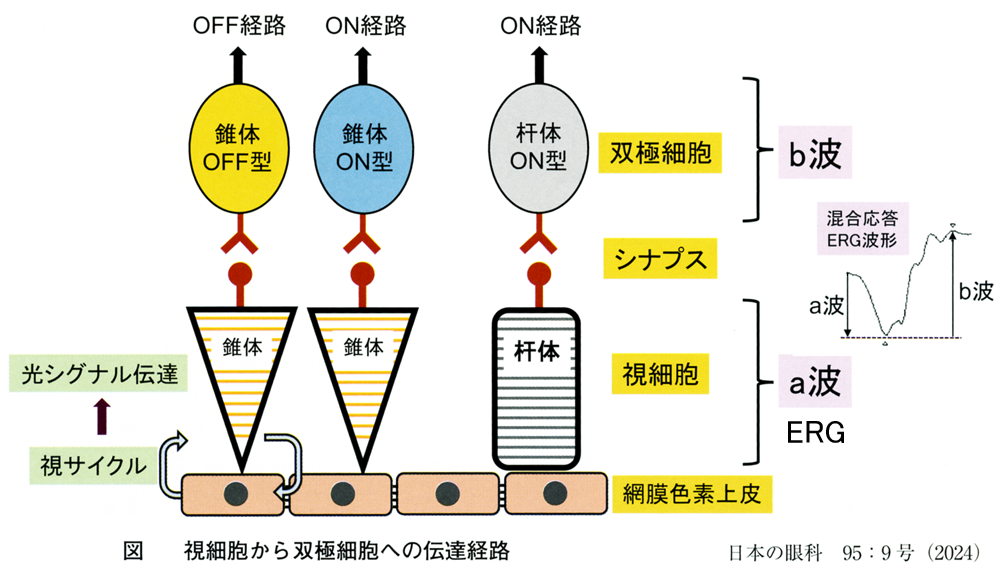
双極細胞は,on型(光増強時,あるいは明るい目標で反応.HBC;hyperpolarizing BC)と off型(光減弱時,あるいは暗い目標で反応.DBC;depolarizing BC)とがあり,視覚情報のON·OFF分解を行う.
錐体からの入力を受ける錐体双極細胞はon型とoff型により高解像処理をしている.
杆体からの入力を受ける杆体双極細胞はon型で信号の増幅処理をしている.これらにより,空間・時間の変化すなわち光刺激のコントラストや増減を検出する(on型が90%以上 ¿ 同数 ?).
10種類程,確認されているとのことである.
➊on中心型:光照射で脱分極する(グルタミン酸が結合すると過分極する・・極性が反転.IPL外側にシナプス.
➋off中心型:光照射で過分極する(グルタミン酸が結合すると脱分極する.IPL内側にシナプス.
形状からはおおきく3種類ある.小さな樹状突起の midget双極細胞,放散した樹状突起の diffuse双極細胞,青on信号専用双極細胞,である.
➀midget双極細胞には大小あり,それぞれmidget神経節細胞に接続する.大型は緑経路,小型は赤経路に対応し,緑信号を多く加算することで黄斑色素によるシフトを補正し,高解像力に関与する.青信号に関しては off型双極細胞が存在.
➁diffuse双極細胞は parasol神経節細胞に接続する.LM錐体が収束し濃淡コントラストに関与する.
➂青のon信号専用の双極細胞は二層性(bistratified)神経節細胞に接続する.ここには赤・緑の和信号がoff型diffuse双極細胞から入力し,青on-黄offの色対立信号となる.
双極細胞はamacrine細胞・神経節細胞とシナプス結合をしている.双極細胞は脱分極によりグルタミン酸が放出され,同じ極性の応答が神経節細胞・amacrine細胞に発生し,情報がリレーされる.
![]() 視覚の本質は,光を検出することに意味がある.多数の杆体からの入力の収斂があるので高い光感度(所謂,
視覚の本質は,光を検出することに意味がある.多数の杆体からの入力の収斂があるので高い光感度(所謂,
☆amacrine細胞:
amacrine細胞は軸索がなく,その樹状突起を横方向に伸ばし,細胞同士の結合と双極細胞や神経節細胞とシナプスを形成している抑制性介在ニューロンで,GABAやグリシンなど(数種類の伝達物質放出)の抑制性伝達物質により側方抑制を行っている.
光照射により持続性の応答を示すものと,光照射の初めと終わりに一過性の脱分極を起こす細胞とがある.時間軸の中で視野の中の変化を検出しているらしい(ぶれの補正,SN比の確保,
・内網状層では視覚の時間的情報処理を行う.側抑制により微小変化を検出する.
・語源は a- (ἀ-) „non“,makros (μακρός) „large“,ines (ἴνες) „fiber“,ということで,かつては無軸索細胞の名があった.なお詳細には,広く軸索を延ばしている多軸索細胞も含むなど,多くの種類があるそうである.
☆網状層間細胞:内網状層と外網状層との間に突起を伸ばしている,amacrine系ドーパミン細胞.
黄斑部には約半数が集中している.神経節細胞は網膜の出力細胞で唯ひとつ,活動電位 action potentialsを発するニューロンである.
網膜神経節細胞は,真っ暗闇でも低頻度でランダムに放電している.これを自発放電という.自発放電のスパイクは光の照射で増減する.網膜神経節細胞の受容野は,中心野と周辺野がある.
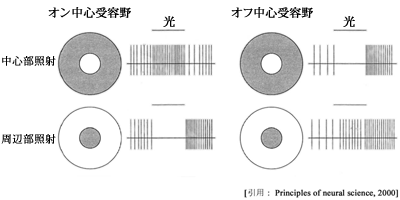
a.神経節細胞は,ON応答しか示さない ON 型神経節細胞,OFF応答しか示さない OFF 型神経節細胞,両方に応答する ON 型 OFF 型神経節細胞が存在する.受容野は,中心部を照射するときと周辺部を照射する時の反応は拮抗的である.
①on型受容野:中心-on,周辺-off 型ニューロン ( on-center,off-surround type )
光照射に対して,中心が興奮性に周辺が抑制性に応答すること.中央部を照射するときパルス発射頻度が増加(発火)し,周辺部を照射すると刺激中は抑制され,off直後に発火する.明るい背景にある暗い点に強く反応する.
②off型受容野:中心-off,周辺-on 型ニューロン ( off-center,on-surround type )
中心が抑制性に周辺が興奮性に応答すること.上と逆の反応を示す.光減弱で発射頻度が増加,暗い背景の中で光る点に強く反応する.
受容野とは,呈示された光刺激に対して,網膜内の細胞が電位応答する空間領域のことである.受容野内では,中心の周辺に対する明るさ(on部分)と暗さ(off部分)のコントラストを伝える信号,あるいは対象とする物体の明るさよりもその輪郭を特徴として抽出するような性質をもつ,といえる.
b.XYW系 【 追加事項 】
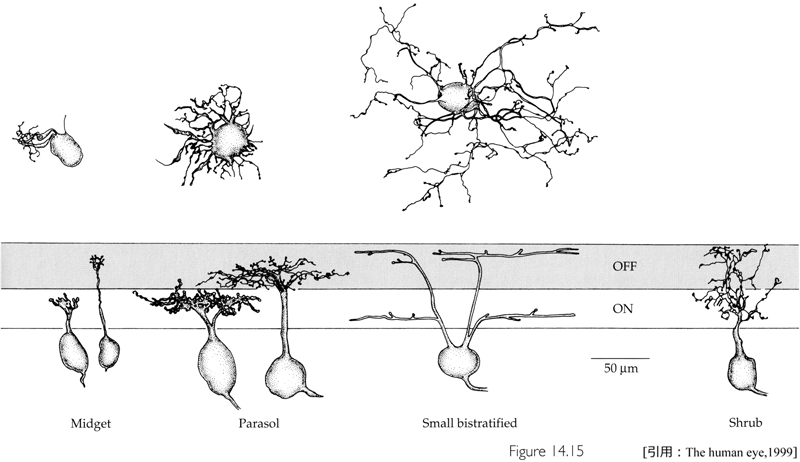
網膜レベルで錐体と杆体の二種類の視細胞に与えられた情報は,網膜神経節細胞に至り色覚・形態覚・コントラストや動きなどの情報として処理される.神経節細胞は機能的・形態学的に独立したいくつかのグループに分けられ,視覚情報の特性(modality)を抽出し複数のチャンネルで信号を出している(並列処理 parallel processing という).代表は小さいX型(midget細胞)と大きいY型(parasol細胞)である.両者で全数の約80%.
❶midget神経節細胞(X細胞または β細胞)は軸索が細く伝導速度の遅い小型の神経節細胞で,光照射に対し一定した持続性
(sustained型)
の応答・線形加重を示す.波長により反応強度は変化する.受容野は小さく,空間周波数感度にすぐれる.時間分解能は低い.
対象となる静止物体の高解像度の像や色覚情報(形態覚 form vision)を担うことで,コントラスト感度では高周波部を受け持つ.
黄斑に分布し,全神経節細胞の約70%.線維はほとんどが外側膝状体 parvo cell群へ投射し,一部が視蓋前域に投射.
❷parasol神経節細胞(Y細胞または α細胞)は軸索が太く伝導速度が速,輝度変化に対し一過性
(transient型)
すなわち敏感に応答する.波長の違いで反応は変わらない.受容野は広く,時間分解能にすぐれる.空間解像度は低い.
視覚対象の位置・奥行きや動きの情報(動態覚 motion vision)を担うことで,コントラスト感度の低周波部を受け持つ.
網膜全体に分布し,全神経節細胞の約10%.線維は外側膝状体 magno cell群と上丘(外側膝状体外視覚系として視床枕を経由し18野へ投射)に接続.
輝度変化への応答ということで,視野測定 とはparasol細胞の機能をみている,と説明される.
❸small bistratified(小型二層性)神経節細胞(W細胞または γ細胞)は中等度の太さの軸索と中等度の伝導速度を持つ神経節細胞である.色覚の一部(blueⲻyellow pathway)と視覚認知以外の眼球運動,瞳孔反射などの視覚性反射に必要な情報を伝える.
全神経節細胞の約10%.線維の大部分は上丘に,一部は外側膝状体 konio cell群に投射する.視蓋前域核に入る情報もある.
・large bistratified ganglion cells
・層 stratificationとは:内網状層は大きく6層に分けられる.神経節細胞は樹状突起が存在する層により,単層性・二層性・多層性に,さらにON型・OFF型に種別される.おおよそ,神経節細胞寄りがON層,内顆粒層寄りがOFF層となる.
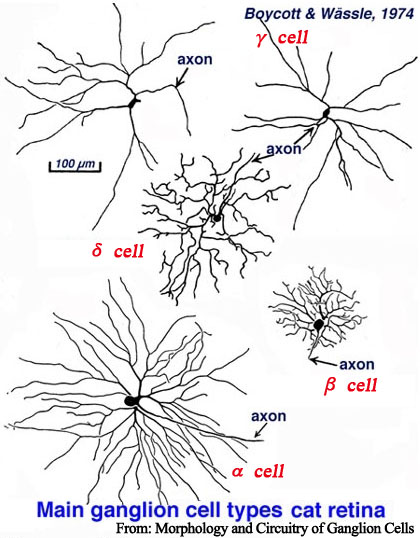
・樹状突起の広がりは:主にネコ網膜の組織学(形態学)的研究により,広いほうから α・β・γ ・・・ と名づけられている.
α細胞:広い領域に疎に張っている.
β細胞:狭い領域に密に張っている.
γ細胞:細胞体がβ細胞より小さく樹状突起が少ないが広い.
δ細胞:細い樹状突起を広く張り出す細胞で,γ細胞の一種(亜型)とされる.
ε細胞:視床枕に投射.
・その他,外側膝状体内の接続先はもとより膝状体外の行き先が決定されていない数種類があるとのことである.
c.メラノプシン神経節細胞(内因性光感受性網膜神経節細胞
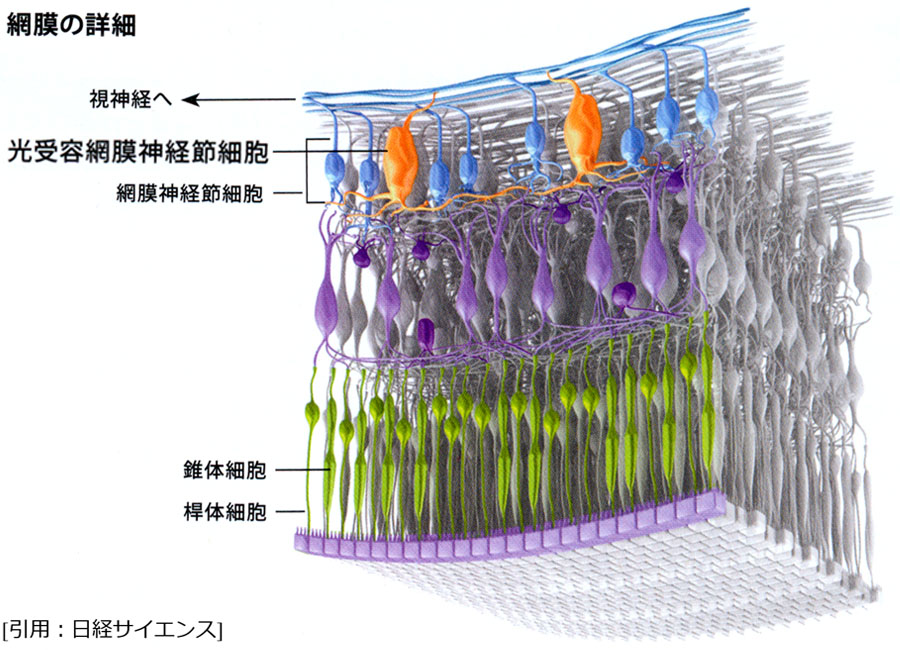 多くの大きな樹状突起を持つ
神経節細胞
で,青色光(400〜500nm)を感受(ピーク470〜485nm)し,
視交叉上核 ・松果体へ接続し,メラトニンの産生抑制・概日周期の調整をしている.一部は視蓋前域上丘オリーブ核へ入力し,外界の光強度を感知し瞳孔反応に関与する(非視覚応答 nonⲻimage forming pathway).すなわち対光反射は二系統の光情報に因る.これらにより,脳内の明るさ情報符号化処理(irradiance encoding process)を担っていると説明される.
多くの大きな樹状突起を持つ
神経節細胞
で,青色光(400〜500nm)を感受(ピーク470〜485nm)し,
視交叉上核 ・松果体へ接続し,メラトニンの産生抑制・概日周期の調整をしている.一部は視蓋前域上丘オリーブ核へ入力し,外界の光強度を感知し瞳孔反応に関与する(非視覚応答 nonⲻimage forming pathway).すなわち対光反射は二系統の光情報に因る.これらにより,脳内の明るさ情報符号化処理(irradiance encoding process)を担っていると説明される.
W型細胞系と共に羞明を惹起する網膜の起点のひとつと目され,短波長と羞明を関連づける根拠となっている.視細胞へフィードバックする信号回路の存在も指摘されている.
視細胞の光受容とは無関係に脱分極(感桿型光受容細胞)を起こすことで発見された.
また,片頭痛発作に関連する光感受性細胞ともみられている.0.2%ほど ?
d.そして 続きは 外側膝状体 で
形態覚や色覚の情報は parvoⲻcellular pathway(Pⲻ系)へ,コントラストや動きの情報などは magnoⲻcellular pathway e.さらに Y・X・W(Y:X:W=1:8:1)はネコ網膜にて生理学的に研究された命名で,軸索伝導速度や受容野,反応特性などから分類された.これらは,α細胞はY細胞に,β細胞はX細胞に,γ細胞はW細胞に,それぞれ対応している.完全にではないが,霊長類(P;primate)とくにサルでの細胞形態にもおおよそが一致するとのことで,parasol細胞を Pα,midget細胞を Pβ ということも多い.が,そもそも敢えて区別せずに記述される教科書も多いようである. 発生途上,神経節細胞の前駆細胞は過剰に作られる.軸索が脳内各所へ届く確率を計算しているのだろうか.7割の前駆細胞が消えるとのことである.さらに生後においても年あたり5,000本の軸索が消えていくのだとか. f.念のため 40種類以上の神経節細胞が確認されているそうである.
すなわち 基本的な反応は
光が光受容器に到達すると視色素が分解される.光化学反応である.これにより視細胞は過分極の状態になり,伝達物質の放出量が減少する(ここまでで光情報→化学情報→電気情報と変換された).
通常,伝達物質は双極細胞を過分極の状態にするため,放出量の減少によって双極細胞は脱分極の状態になる(ここで視細胞→双極細胞に情報が伝達された).
双極細胞は脱分極によって伝達物質を放出し,神経節細胞を興奮させる(活動電位発生 ・・・ ここで視神経に視覚情報が乗ったことになる).
 |
 |
このようにして符号化・インパルスとなった情報は神経節細胞から発信され,視神経により
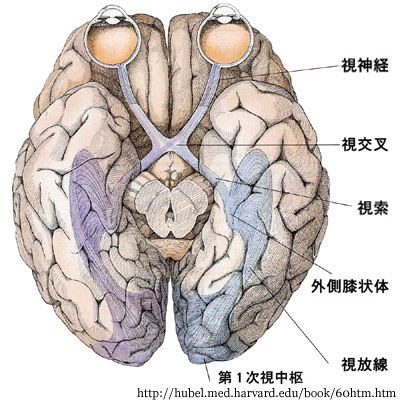
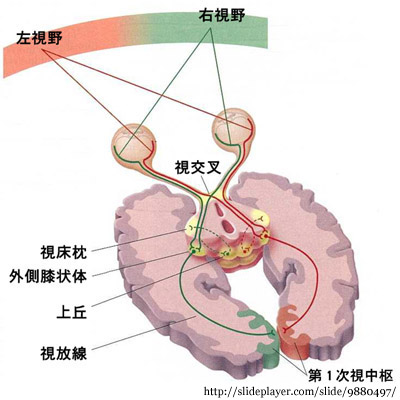
主として視床下部の外側膝状体に,一部は上丘等に伝えられる( →
【第九章】 視 路 へ).
![]()
●細胞膜電位(分極)と Na-K ポンプ 【 膜電位 】
細胞の環境は,外液の Na+・Ca2+・Cl-,内液の K+,の濃度勾配がある.細胞膜において Na-K ポンプとよばれる能動輸送系により,外に Na+,内に K+のイオン勾配が維持されている.すなわち, Na+濃度は 内<外, K+濃度は 内>外,となっている.静止時には細胞膜のカリウムチャネルのK+透過性によりK+が外に流出しようとすることで,細胞膜を境として,細胞の内側は-(マイナス)に外側は+(プラス)となっている.これを分極している,という.静止膜電位は,通常(細胞外をゼロ電位として)−70mV〜−60mV に保たれている.一般に神経細胞の活動では細胞膜電位は一過性にプラス側に振れる.スパイクである.よーするにインパルスという活動電位で情報が伝達される.ただし,光受容時の視細胞の反応は電位変動による.

●脱分極(depolarization):膜電位が上がること.膜電位を上げるのに影響するイオンは Na+・Ca2+.活動時に Na+が細胞内に流入することによる.
視細胞内節では細胞内から細胞外へ Na+を吐き出している(Na-Kポンプ).平行して細胞外K+が流入するが同時に選択的K+チャンネルにより細胞内K+が流出しバランスが保たれている.他方,外節では陽イオンチャンネル(cation channel:cGMP依存性 Na channel:Naチャンネル)が開いた状態で,形質膜を介して細胞内に陽イオン(主として Na+,と Ca2+)が流入している(dark current:暗電流).これにより外節の膜電位はおよそ −40〜−30mVであり,暗順応時は脱分極状態にある,と説明される.cGMP濃度により Naチャンネルが開閉することで,cGMP依存性という.
●過分極(hyperpolarization):膜電位が下がること.膜電位を下げるのに影響するイオンは一般に K+,
視細胞では rhodopsinが光エネルギー粒子(フォトン photon)を吸収し,11-cis型retinal を 全trans型retinal に変換すると(異性化),オプシンが活性化する.
→ 中間産物メタロドプシン が トランスデューシン(tansducin:興奮性のG蛋白)を活性化する.
→ tansducin は フォスフォジエステラーゼ(phosphodiestelase:PDE )を活性化する.
→ PDE は cyclic guanosine monophosphate(c-GMP:環状グアノシン一リン酸)を guanosine monophosphate(5'-GMP
→ cGMP濃度がさがることで Na+channel が閉鎖(流入停止と,Na-Kポンプによる細胞内 Na+の流出が続き細胞内の電位はさらにマイナスになる)
→ 膜電位が下がる,すなわち過分極化(ERGa波の発生).
このとき視細胞へは K+が取り込まれ続け,細胞外 K+は減少する.一方で内顆粒層での細胞外K+は増加する.amacrine細胞と神経節細胞,双極細胞の脱分極によるものである.この反応は強く長時間つづき,この増加分の K+は Müller細胞に取り込まれる.
●Müller細胞
基底部(基底膜)は硝子体に面して内境界膜を,核は内顆粒層,頂部/突起部は視細胞部で外境界膜を形成する.さらに多数の細突起を隣接する神経要素~視細胞間/網膜下腔へ伸ばしている.
網膜血管に対しては,血液関門の一役をになう.毛細血管基底膜の一部にもなっている.細胞外液の成分を調節する.普段はGDNF(glial-cell-line-derived neurotrophic factor)を産生して血管透過性を下げているが,糖尿病ではVEGFを産生して透過性亢進をきたす. また,錐体視物質(vit A)のリサイクルを担うといわれる.
変性・壊死などの神経細胞の脱落があると増殖して,損傷を受けた網膜色素上皮を含む網膜欠損部や閉塞血管内腔の穴埋めをする.網膜外(特に硝子体中)に移動・増殖して増殖膜の成分にもなる.
感覚網膜の発生時には他の神経細胞よりも早く最初に前駆細胞(神経膠芽細胞)から細胞的分化を開始する.胎生期にはヒアルロン酸を産生する.
これらにより網膜を構造的・機能的に支える.最近の研究では神経細胞に再分化する潜在機能(リプログラミング)が証明され,網膜再生への手がかりが期待されている.
グリア細胞の細胞膜透過性はおもに K+であり,静止膜電位は細胞外液の K+濃度に依存する.視細胞過分極による視細胞腔(細胞外)のK+濃度が上昇すると細胞内に流入し,内境界膜から硝子体中へ吐き出す.この移動により ERGb波が生じる.
⏩ 生体の光ファイバoptical fibers:light guide
●オプシン【opsins】
様々な生物における光受容分子で,光利用の基となるタンパクである.
機能的には「視覚系オプシン visual(image᠆forming) opsin」と「非視覚系オプシン nonvisual(non image᠆forming) opsin」,
分布・構造により,「微生物型オプシン typeⅠ opsins 」と「動物型オプシン typeⅡ opsins 」に分けられる.
我々の視物質は繊毛性光受容体細胞で発現するサブタイプ・繊毛オプシン(ciliary opsins)に属する.
| 名称 | 略称 | 最大吸収 | 色 | 網膜分布 |
| 赤錐体(L-cone) | OPN1LW | 560 nm | 黄 | 錐体 |
| 緑錐体(M-cone) | OPN1MW | 530 nm | 緑 | 錐体 |
| 青錐体(S-cone) | OPN1SW | 430 nm | 紫(violet) | 錐体 |
| ロドプシン | OPN2(RHO) | 500 nm | 青-緑 | 杆体 |
| エンセファロプシン (パノプシン) | OPN3 | (470nm) | 青-緑 | 杆体 錐体 外網状層 内網状層 神経節細胞層 |
| メラノプシン | OPN4 | 480 nm | 青 | 内因性光感受性網膜神経節細胞:ipRGC (intrinsically photosensitive retinal ganglion cell |
| ニューロプシン (オプシン 5) | OPN5 | 380 nm | 紫外~紫 | 感覚網膜 網膜色素上皮細胞 |
| ペロプシン | RRH | 色素上皮微絨毛 (retinal pigment epithelium-derived rhodopsin homolog | ||
| Gタンパク共役型受容体 | RGR | Müller細胞 網膜色素上皮細胞 |
●固視微動
神経細胞は変化分にしか応答しない.よって,静止画をじっと見ていると(動きがないと)細胞は何も応答しなくなることになる.実際には眼球運動を随意的に止めることはほぼ無理であるばかりか,生理的に常に細かい動きがある.したがって輪郭・境界の位置も細かく変化し,常に信号が出ていることになる.固視微動である.
神経節細胞の易疲労性とか,皮質レベルでの神経細胞反応特性による,
●側抑制:変化の検出
与えられた光刺激は,光を受けた神経節細胞を興奮させると同時に隣接した細胞を抑制するようになっている.信号の始まる時にだけあるいは終わる時にだけといった,信号が時間的に変化した時に強く反応するのは、時間軸上の側抑制の機能となる.側抑制によって,明暗あるいは色彩が変化する境界で反応が現われる.色については灰色から赤(緑),青(黄)に変化することを検出する.
側抑制によると,一様な明るさの部分では神経節細胞はほとんど反応しないことになる.つまり本当に暗い所でも,非常に明るいところでも、そこが一様に明るいとやはり細胞はほとんど反応しない.明暗判断は見ている部分の明るさではなく,視野全体での相対的な明暗の変化で行う.
反応が現われる所は明暗あるいは色彩が変化する境界で,変化分だけ検出して伝送し,後で元の情報を復元している.
●視覚受容野:receptive field
あるニューロンを活動させ得る光刺激空間の範囲.一般に,感覚系の一個のニューロンが受け持つ感覚刺激の空間的な範囲を受容野という.
通常,網膜中心は一対一か狭い範囲で,周辺は受け持ち領域が広い.
受容野からの信号は一個の神経節細胞に集合する.寄せ集め である.
●輪郭強調あるいは辺縁対比:border contrast
網膜に写った像をシャープにする効果(エッジ強調)は神経節細胞の受容野の構造(center-surround構造),すなわち相反的(拮抗的)な同心円状の受容野に起因しているものと思われる.
オン中心・オフ周辺型ニューロンは,受容野中心部の視細胞から興奮性の影響を,周辺部から抑制性の影響を受けていると考えられる.
明暗の境でのオン中心・オフ周辺型ニューロンの動きは ・・・
a)明るい光があたっている部位の細胞は興奮性入力と共に抑制性入力を受け,中等度の放電をしている.
b)境界近くで明るい所にある細胞は,抑制性の入力が半分しか入らず,⒜の場合に比較して発火頻度が大きい.
c)境近くで暗い部位のニューロンは明るい部分からの抑制入力のみ受け,他の暗い部分に比較して発火頻度が小となる.
従って明暗の境では,明るい所はより明るく,暗い所はより暗いように強調される(Mach効果).